Partager la publication "Variations morphogénétiques de « polydactylie » et de « polyphalangie » : une « contradiction évolutive » flagrante"
Par Timmermann Claude
Variations morphogénétiques de « polydactylie1 » et de « polyphalangie » : une « contradiction évolutive » flagrante.
Résumé : On verra ici, sur l’exemple du nombre des doigts ou des phalanges, comment la théorie « saltationniste » (promue par Gould pour expliquer l’absence des fameux « chaînons manquants »), s’expose à la même critique que le gradualisme darwinien : on peut tout y démontrer et aussi bien le contraire de tout2. Posant d’abord qu’il n’y a pas de « retour en arrière », on en déduit que les Mammifères auront tout au plus 5 doigts (qu’ils peuvent toutefois « perdre » pour passer à 3 ou 2). Or il en existe ayant 6 doigts, comme l’Éléphant et le Panda. De plus, les deux espèces de Panda, clairement distinctes, ont toutes deux cet organe surnuméraire : on parlera alors de « convergence adaptative ». Ce n’est là qu’une contorsion verbale montrant bien que le darwinisme est une théorie purement « langagière » (René Thom) : on invente des concepts sur mesure pour faire entrer la réalité dans un cadre préétabli.
L’évolution peut se résumer dans la formule énoncée par le paléontologue Louis Dollo3 en 1893 et reprise par Stephen Jay Gould en 1970 : « L’évolution est discontinue, irréversible et limitée dans le temps. »
Elle s’énonce en ces trois propositions :
I – L’évolution est saltatoire, se produisant par sauts brusques ;
II – Un organisme ne peut revenir, même partiellement, à un stade antérieur déjà réalisé dans sa série ancestrale ;
III – Tous les organismes s’éteignent nécessairement, au terme d’un cycle évolutif qui peut être extrêmement long.
Tout le travail des évolutionnistes concourt à faire entrer dans ce « moule » la multiplicité des organismes fossiles ou vivants, connus et analysés.
Pour ce qui est des membres, chez les Vertébrés Tétrapodes, on se réfère, comme premier type « sorti des eaux », à la lignée des Icthyostégidés dont le membre dériverait des nageoires paires (pectorales et pelviennes) d’un poisson Sarcoptérigien, du groupe des Rhipidistiens : Eustanoptéron (du Dévonien).
Cette hypothèse provient surtout de ce que les Sarcoptérigiens ont des nageoires reliées à une ceinture pectorale (ou pelvienne) bien individualisée, et articulées sur la base d’un seul os (baptisé alors humérus ou fémur), et enfin disposent d’une respiration double branchiale et pulmonée « primitive » de type sacculaire…
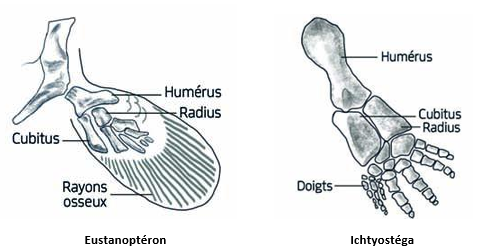
L’idée suggérée ensuite est, qu’au fil de l’évolution, les doigts vont se limiter en nombre, des Amphibiens aux Reptiles, petit à petit à 5, voire moins, et vont perdre des phalanges jusqu’à 3 ou 2.
L’anatomie comparée du membre des Mammifères conduit à faire admettre alors une unité morphogénétique du membre dit « chiridien », fondamentalement donné alors comme pentadactyle, dont tous les types observés vont dériver par hypertrophie, fusion ou perte de certains de ses éléments.
On cherchera donc à faire coller ce modèle de membre aux structures osseuses observées, limitées par suite de « pertes évolutives » à cinq doigts, chacun à 2 (pouce) ou 3 (phalanges), selon le schéma ci-dessous :
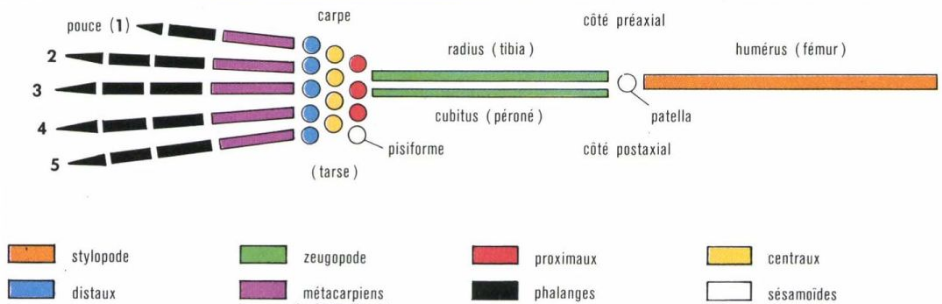
Tout le travail actuel des évolutionnistes concourt à faire entrer dans ce « moule » les membres de tous les Tétrapodes connus pour justifier la théorie.
En particulier chez les Mammifères, on cherchera systématiquement à faire coller le modèle aux structures, notamment osseuses, observées.
Mais la difficulté parait insurmontable lorsqu’on se trouve en présence de structures manifestement surnuméraires, car cela contrevient au dogme : en vertu de la seconde « loi », précédemment énonceée, il est évident qu’un organe « perdu » à un stade donné de l’évolution ne peut « réapparaître » dans un stade ultérieur…
C’est précisément cette contradiction qui est illustrée par l’hyperdactylie et par l’hyperphalangie…
(Nous parlons bien ici de standard d’espèce et non pas de phénomènes plus ou moins tératologiques, bien connus, tels chez l’Homme les sexadigitaires… assez nombreux en Afrique noire.)
Polyphalangie
C’est le cas de certains cétacés du nouvel ordre évolutionniste des «Cétartiodactyles » qui regroupe :
- l’ordre des Artiodactyles (Ongulés à nombre pair de doigts, dit aussi Paraxoniens, ou Paradigités)
- l’ordre des Cétacés (considéré maintenant comme « étant secondairement retourné à la mer »)
Et là certaines espèces (pas toutes) ne sont pas du tout dogmatiquement correctes, en développant chez les Mysticètes (Cétacés à fanons) tels la Jubarte ou Mégaptère, comme chez les Odontocètes (Cétacés à dents) tels certains dauphins, une polyphalangie parfaitement incongrue aux regards de la norme.

Notons cependant que les Cachalots ont bien appris leur cours d’évolution et présentent des membres antérieurs corrects à cinq doigts ayant le nombre de phalanges requis…
De même les Siréniens (Lamantins) et les Pinnipèdes (Phoques) ont tous des membres antérieurs parfaitement conformes au modèle, n’ayant sans doute pas la nostalgie d’ancêtres galopant dans les grandes plaines…
« Polydactylie »
La polydactylie vraie apparaît en revanche toujours anormale4, pathologique, mais peut se transmettre dans certains cas génétiquement dans des lignées animales, comme dans des familles humaines (notamment les sexadigitaires), ce qui montre que le « pool génique » dans de nombreuses espèces n’exclut pas cette réalisation et sa transmission.
À l’inverse, certains animaux ont systématiquement développé de « faux doigts » pour compléter leur métacarpe ou leur métatarse en lien avec une fonction précise : un véritable « paradoxe évolutif ».
Le « sixième doigt » de l’éléphant
Les doigts de l’éléphant ne sont pas tous terminés par un ongle apparent, ce qui crée déjà confusion !
Ainsi, l’éléphant d’Afrique en possède 3 sur ses pattes arrière contre 4 pour l’éléphant d’Asie.
Sur les pattes avant, l’éléphant d’Afrique présente 4 ou 5 ongles contre toujours 5 pour l’éléphant d’Asie.
Les 5 doigts de la patte sont orientés plutôt vers l’avant et reposent sur un coussin fibreux conique, donnant l’impression que le pied est planté sur ses orteils. Ce coussin joue le rôle d’amortisseur et aussi de récepteur des vibrations du sol, que l’animal perçoit de très loin.
Les phalanges sont quasiment soudées et ne permettent pas de latitude de mouvements aux doigts, noyés dans la masse du pied, qui ne servent qu’à équilibrer le métatarse et le tarse, calés sur le coussinet plantaire qui sert ainsi d’assise…
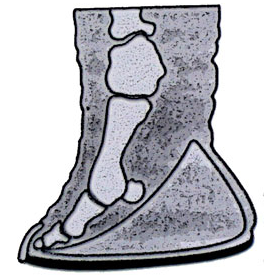
On constate en arrière d’un métacarpe (ou d’un métatarse) la présence d’une excroissance osseuse improprement baptisée « sixième doigt » .
Il s’agit en fait de l’os sésamoïde (os de membrane, non squelettique), qui s’est hypertrophié et qui participe à l’assise du pied sur le coussinet plantaire en y pénétrant profondément (long de 5 à 8 cm), où il agit comme transmetteur direct des vibrations du sol, réceptionnées par le coussinet fibreux.
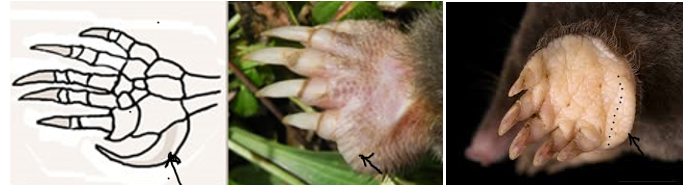
Le faux « pouce » du panda
La modification d’un os sésamoïde se retrouve aussi chez d’autres espèces dont l’exemple le plus célèbre est le fameux sixième doigt du panda.
Il s’agit du résultat du développement de l’os sésamoïde radial (R S) (cf. figure ci-dessous) allongé en doigt, qui s’articule pour former un faux pouce opposable, terminé par une grosse pelote digitale qui le recouvre.
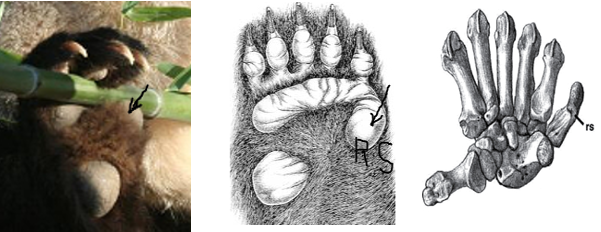
La « pince digitale » ainsi formée a été largement évoquée par les évolutionnistes pour illustrer l’adaptation remarquable que constitue un tel dispositif préhensile en lien avec la nutrition très particulière de l’animal : la monophagie de la tige de bambou.
En fait, cette propriété n’est pas spécifique de l’espèce, mais se retrouve chez les deux espèces de pandas.

Le « grand panda » aussi appelé « panda géant » (Ailuropoda melanoleuca) précédemment évoqué, classé parmi les Ursidés, et le « petit panda » aussi connu sous le nom de « panda roux » (Ailurus fulgens) classé parmi les Procyonidés (Ratons laveurs) où on lui retrouverait un ancêtre (?) fossile le Simocyon : Simocyon batalleri, frugivore et carnivore arboricole du Miocène d’Espagne, présentant la même particularité anatomique du « faux pouce5 ». Les deux pandas ne sont donc pas directement voisins en terme d’espèce, mêmes s’ils partagent le même habitat et sont endémiques des mêmes forêts chinoises du Tibet et de certaines autres zones forestières himalayennes.
L’un restant terrestre et l’autre arboricole.
La présence de ce « faux pouce » a donc été l’occasion d’une débauche de commentaires et de publications sur la « convergence adaptative » observée entre les deux espèces.
Le grand panda est volumineux et massif : il pèse de 80 à 125 kg, avec une moyenne de 105,5 kg ; il mesure de 1,50 à 1,80 mètre de longueur, avec une moyenne de 1,65 mètre.

Le petit panda (figure ci-dessus) est de la taille d’un renard : le corps sans la queue mesure de 50 à 65 cm, la queue mesure de 30 à 60 cm. Les mâles pèsent de 3,5 à 6,2 kg et les femelles de 3 à 6 kg.
Ils sont arboricoles et omnivores à prédominance frugivore, mais sans négliger les petits rongeurs, les œufs et les oisillons qui pourraient tomber sous leur patte… et bien entendu le bambou…
L’appendice falciforme de la main de la taupe (Talpa europaea)
La taupe (Talpa europaea) présente également un « sixième doigt » ennoyé dans l’épiderme granuleux de la patte antérieure.
Il s’agit là encore du sésamoïde radial qui se développe sous forme d’un élément osseux falciforme qui vient élargir la paume du métacarpe et intervient en renfort sur la tranche, côté radial, sans pourtant apparaître extériorisé dans le membre.

On comprendra sans peine que ces deux fonctions contribuent largement à renforcer l’efficacité de l’action de la patte lors du fouissage qui peut atteindre en longueur de galerie plus de 80 cm par jour !
Mais on notera qu’il existe dans la même niche écologique un Rongeur sensiblement de même taille et ayant le même comportement : le campagnol terrestre ou rat taupier (Arvicola terrestris) aux performances de tunnelier tout à fait comparables à celles de la taupe.
L’analyse qui consiste à dire que la « pelle » formée par le rajout de cet os falciforme à la structure de la main de la taupe serait une « adaptation au fouissage » – ce qui est bien sûr immédiatement mis en avant par les évolutionnistes darwiniens – est évidemment totalement abusive : les performances comparables du rat taupier le démontrent….
Conclusion
Les exemples évoqués montrent soit le renforcement de la palette natatoire par hyperphalangie, soit un renforcement du squelette de la main par le développement d’un os de membrane : le sésamoïde.
On constate embryologiquement que la croissance de cet os intervient secondairement après la mise en place définitive du membre et ne procède donc pas de l’architecture du squelette à proprement parler.
L’existence de cet os est donc liée à sa caractéristique fonctionnelle, ce dans quoi s’engouffrent le concept d’adaptation et la philosophie qui s’y rattache.
Ces exemples montrent clairement les limites de la conceptualisation évolutionniste d’essence darwinienne, et mettent en relief sa contradiction intrinsèque.
L’évolution, chez les Mammifères, aurait conduit à la simplification d’un membre déjà devenu pentadactyle – triphalangé (deux seulement pour le pouce) – ne pouvant plus se transformer qu’à la faveur de la modification, ou de la délétion, de certains de ses éléments.
Pourtant, à l’encontre de ce schéma, même en les acceptant pour la démonstration, on constate alors qu’à partir d’un certain stade l’exigence de fonctionnalité du membre (et ses performances), dans diverses espèces, a imposé de pallier certaines des délétions évolutives, déjà advenues, par l’adjonction nouvelle d’éléments supplémentaires, justifiés au nom de l’adaptation,dont la présence même va à l’encontre des lois de l’évolution !
1 Nous entendrons ici, par extension, « polydactylie » comme phénomène d’existence constatée de pseudo-doigts, ou de structures avoisinantes, surnuméraires par rapport à la « norme évolutive » établie : le membre pentadactyle.
2 Ndlr. En termes savants, nous pourrions dire que, dans une théorie contradictoire, toute proposition (tout énoncé, aussi bien l’énoncé A que son contraire non-A) est un théorème (peut être démontré à partir des postulats).
3 L’ingénieur qui découvrit les fameux ignanodons de Bernissart (en Wallonie, entre Valenciennes et Ath) et leur consacra le reste de sa vie.
4 On pourra consulter avec profit les documents suivants (publiés sur internet)
- Thèse de Dr vétérinaire de Julie VALLOIRE-LUCOT,
- « LA POLYDACTYLIE CHEZ LES MAMMIFÈRES », 2012.
http://theses.vet-alfort.fr/telecharger.php?id=1454
- Louis BLANC, art. « Étude de la Polydactylie chez les mammifères » , Publications de la Société Linnéenne de Lyon, année 1893, 40, p. 53-88.
( http://persee.fr/doc/linly_1160-6398_1893_num_40_1_4046 )
5 SALESA M. J., ANTON M., PEIGNÉ S. & MORALES J. (2006), « Evidence of a false thumb in a fossil carnivore clarifies the evolution of pandas”, Proceedings of the National Academy of Sciences, 103, (2), 379-382.
SALESA M. J., ANTON M., PEIGNÉ S. & MORALES J. (2008), “Functional anatomy and biomechanics of the postcranial skeleton of Simocyon batalleri (Viret, 1929) (Carnivora, Ailuridae) from the Late Miocene of Spain”, Zoological Journal of the Linnean Society, 152(3), 593-621.