Partager la publication "Des yeux pas comme les autres"
Par Primavère, Bruno
Résumé : Darwin avouait, de l’œil, qu’il lui donnait la fièvre. Devant cet organe complexe finement relié à des zones cérébrales qui, en « interprétant » les images optiques, nous permettent de « voir », comment croire en effet que cet ensemble merveilleusement coordonné se soit formé peu à peu sous l’effet du hasard ? Qu’appporterait un œil incomplet, donc non fonctionnel, dans la lutte pour la vie ? Au fond, d’ailleurs, qu’est-ce que la « vision », cette faculté de se représenter le monde extérieur, de s’y orienter, d’y apprécier les couleurs et les distances ? Le mystère ne demeure-t-il pas entier ? Or les outils modernes d’investigation nous montrent au sein du règne animal une diversité d’yeux et de procédés visuels, qui établit clairement comment chaque espèce a reçu du Créateur un ensemble d’organes préadaptés à la vie qu’elle était conçue pour mener : vie nocturne ou diurne, aérienne ou aquatique, etc. Aujourd’hui, ce n’est donc plus une banale fièvre qui menace les darwiniens, mais l’écroulement définitif de leur croyance.
Claude Destaing1 et les Drs Louis et Paul Murat2 ont présenté l’œil et sa complexité dans deux livres certes anciens mais qui n’ont rien perdu de leur actualité.
Toutefois, les moyens d’investigation dont nous disposons ne cessent d’amplifier les connaissances et cette complexité s’en trouve mieux mesurée.
En physiologie du système nerveux, mentionnons le réflexe vestibulo-oculaire qui permet la stabilisation du regard lors des mouvements de la tête
Il est facile de montrer comment l’appareil vestibulaire participe à la stabilisation du regard lors des mouvements de la tête : regardez un objet face à vous et, tout en continuant à le regarder, tournez rapidement la tête. Le regard ne semblera pas avoir quitté l’objet. Lorsque la tête bouge, les canaux semi-circulaires du vestibule de l’oreille interne signalent à quelle vitesse se fait le mouvement céphalique. Le système oculomoteur répond aux mouvements de la tête par un mouvement des yeux égal en amplitude angulaire mais opposé en direction au mouvement de la tête, et ceci selon les trois degrés de liberté́.
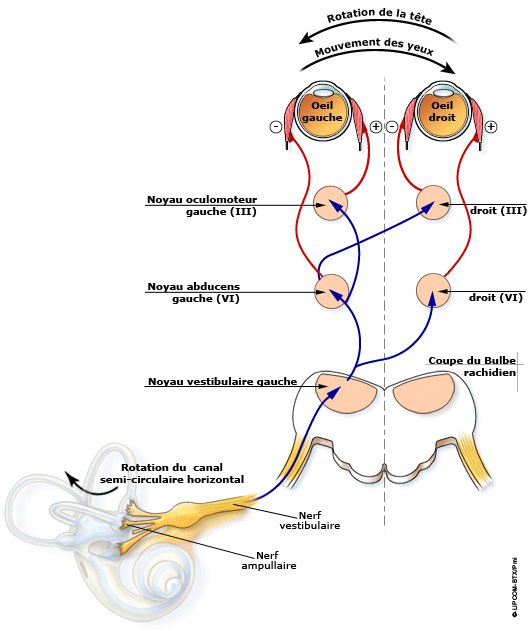
Fig. 1. Coordonner la vision avec les mouvements de la tête. Pas si simple !
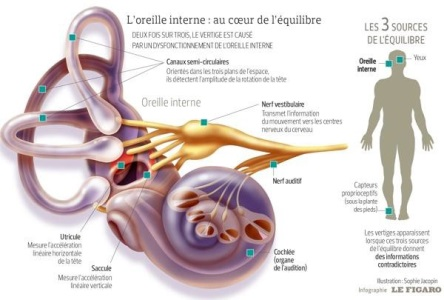
Fig. 2. L’oreille interne, capteur essentiel à notre équilibre.
Après nous être émerveillés qu’une cellule œuf parvienne à donner un corps adulte avec des régionalisations cérébrales, des organes interconnectés selon différentes modalités (nerveuse, sanguine, hormonale), il suffira de nous arrêter sur un composant de notre système visuel : le cristallin, structure d’origine épithéliale (la couche cellulaire concernée pendant l’embryogenèse du cristallin est l’ectoderme, qui donne également la peau, les poils, etc.). Il est formé au cours du développement embryologique par une couche de cellules qui s’invagine (se creuse en se refermant sur elle-même), et forme alors une sphère cristallinienne initialement creuse. Les cellules épithéliales situées au niveau de la face postérieure se différencient et s’allongent, en formant les premières fibres cristalliniennes. Leur croissance comble progressivement le vide central de la sphère cristallinienne, et les fibres du cristallin viennent alors au contact des cellules épithéliales antérieures. Ces dernières vont assurer la croissance ultérieure du cristallin. Il existe en effet une zone située juste à l’avant de l’équateur, sous la capsule cristallinienne, où des cellules épithéliales germinatives se multiplient, et croissent en s’allongeant à l’avant et vers l’arrière, en recouvrant des fibres plus anciennes…
En biologie moléculaire et génétique, les cristallines3 suffisent à nous interpeller : elles se distinguent par leur solubilité dans l’eau. Extrêmement spécifiques, nous ne les trouvons dans aucun autre organe de notre corps bien que toutes les cellules aient la même information génétique. La fabrication d’une seule protéine fait appel à toute une machinerie composée d’ARN (acides ribonucléiques) et de protéines, qui suffit à elle seule, pour l’homme honnête, à infirmer le côté hasardeux et aléatoire de notre appareil visuel. Cette affirmation reste scientifique car elle est une induction a posteriori qui procède de la logique de l’analogie (avec notre expérience, les « créations » artistiques). Une telle machinerie, une intelligence l’a conçue.
Notre raison est conduite à croire en l’existence d’une Pensée créatrice ou au moins d’un dessein intelligent dans l’élaboration du système visuel ; elle est même « acculée » à cela, bien qu’elle demeure libre. Notre intelligence peut, comme devant un ouvrage de l’art humain, refuser l’évidence.


Fig. 3. Une horloge astronomique est-elle le fruit du hasard ou l’œuvre d’une intelligence ?
Elle serait comme cet homme qui, émerveillé par les horloges astronomiques, ne conclurait pas à l’existence d’un concepteur, concepteur dont les dons apparaîtraient d’autant mieux, à mesure que les mécanismes de l’horloge seraient analysés et étudiés.
Cet insensé n’aurait que cette seule perspective : considérer quelle horloge est plus proche de telle autre pour déterminer un ancêtre commun d’horloge (horloge qui présenterait des caractéristiques communes et dont auraient dérivé toutes les autres). Admettre quelqu’un au-dessus de lui contrarierait-il l’orgueil humain ? Ou bien cela amènerait-t-il l’orgueilleux à devoir reconsidérer sa vie et peut-être sa vie morale ?
La démarche qui vient d’être décrite (et qui se réfère à la phylogénie) « pollue » régulièrement le discours scientifique, que ce soit dans les encyclopédies ou les revues de vulgarisation. C’est une manière de « botter en touche » pour écarter Dieu de notre horizon dès qu’un questionnement survient face au mystère. Abusivement, certains pensent pouvoir par là se prévaloir d’un discours purement scientifique alors que leurs interprétations matérialistes sont déjà de l’ordre de la philosophie ou de l’idéologie4. A contrario, la description de l’appareil visuel suffit à elle seule à induire ou à déduire qu’il y a intention et conception intelligentes, qu’il y a une finalité dans l’être vivant qui ne peut provenir du hasard, mais vient d’un être pensant, d’une intelligence. L’existence de Dieu est une vérité à laquelle peut atteindre la raison naturelle. La Révélation, en accord avec la raison, nous dit que cette pensée est Dieu-Amour qui, par la Création, donne l’être et la vie.
Cet appareil visuel, qui se retrouve chez une grande partie des êtres vivants, peut-il être une preuve du transformisme par cette homologie ?
Comme nous allons le voir, l’étude fine de l’œil et de ses annexes donnent de nombreux arguments faisant obstacle à l’évolutionnisme.
Plans d’organisations et relations entre les organismes et leur milieu
Le fonctionnement des organismes repose sur les mêmes grandes fonctions, réalisées par des structures différentes ou non selon les plans d’organisation, dans des milieux différents.

Fig. 4. Les yeux d’une abeille.
Les animaux possèdent des organes photorécepteurs leur permettant de percevoir des images. Une simple observation montre une différence d’aspect entre les yeux des arthropodes (yeux composés) et ceux des vertébrés. Comment les objets sont-ils transformés en image cérébrale avec des yeux qui paraissent si différents ? Par ailleurs, l’eauabsorbeunepartiedesrayonslumineux ;lemilieuaquatiqueestdonc,engénéral,moinslumineuxquele milieu terrestre. De plus, l’eau possède un indice de réfraction de la lumière (n = 1,33) plus élevé que l’air(n = 1). Comment fonctionne chacun de ces types d’organes visuels en milieu terrestre d’une part, et aquatique d’autre part ? Comment le milieu influence-t-il les modalités de la vision ?
Voir dans l’air ou dans l’eau
a) L’œil des vertébrés
L’œil des vertébrés a la forme d’une coupe remplie d’un liquide transparent (appelé humeur) ; il est recouvert de la cornée, transparente. Un cristallin, constitué de cellules vides totalement transparentes fait office de lentille convergente ; il permet la formation d’une image sur la rétine, formée de plusieurs couches de neurones photorécepteurs, qui occupe le fond de l’œil.
Chez la souris (et bien d’autres animaux), l’œil est protégé du milieu aérien desséchant par deux paupières [Fig. 5] et des glandes lacrymales associées.
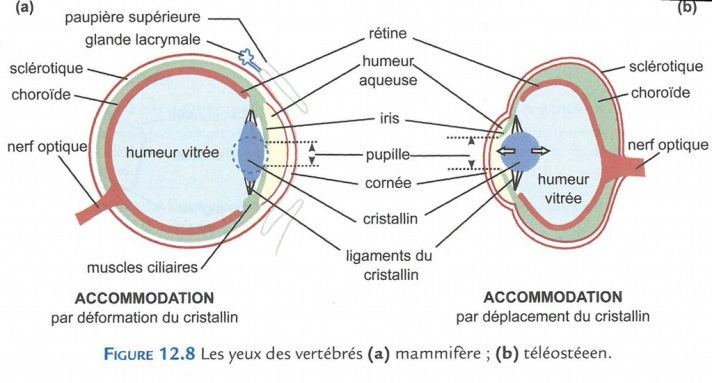
Fig. 5. L’œil d’un mammifère (a) et celui d’un téléostéen (b). L’accomodation se fait par déformation ou par déplacement du critallin.
Derrière la cornée, se trouve l’iris, percé en son centre d’un orifice, la pupille, par lequel la lumière pénètre dans l’œil. Si la luminosité est forte, le diamètre de la pupille rétrécit, ajustant la quantité de lumière qui va frapper la rétine, à la manière du diaphragme sur un appareil photographique. À l’inverse, le diamètre augmente si la luminosité baisse. L’œil des mammifères est sphérique et la cornée bombée ; le cristallin biconvexe réfracte les rayons lumineux issus d’un objet situé à l’infini, de façon à ce que l’image se forme sur la rétine. Pour les objets plus proches, le cristallin se déforme suite à la contraction de muscles attachés aux ligaments du cristallin, ce qui le rend plus convergent et permet ainsi aux images des objets proches d’être formées sur la rétine : c’est l’accommodation. Les rayons lumineux qui frappent les photorécepteurs de la rétine, en un point variable selon l’objet émetteur, déclenchent une réaction photochimique qui, par une cascade de réactions, est convertie en variation du potentiel membranaire des cellules photoréceptrices, elles-mêmes à l’origine d’un message nerveux.
Le traitement par les centres nerveux de cette information permet alors à l’objet d’être vu de façon nette.
Chez les téléostéens (poissons osseux), les yeux sont dépourvus de paupières ; c’est l’eau de l’environnement qui humidifie la cornée. L’œil est aplati selon un axe antéro-postérieur, la cornée est plate, et le cristallin sphérique. Un tel œil permet au poisson de voir nettement les objets immergés ; mais, compte tenu de la différence d’indice entre l’eau et l’air, si cet œil fonctionnait dans l’air, l’image se formerait en avant de la rétine et le poisson serait myope. L’accommodation se fait d’une façon différente de celle des mammifères, par le déplacement du cristallin dans l’axe antéropostérieur de l’œil (un peu comme la lentille d’un appareil photographique lors de la mise au point).
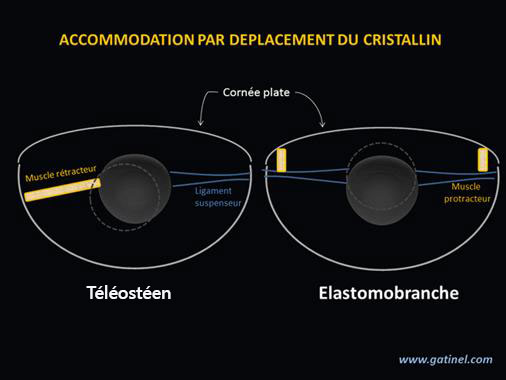
Fig. 6. Muscles et ligaments sont disposés différemment.
Vivre constamment dans l’eau ne veut pas pour autant dire que le reste de l’œil sera semblable. Comme le montre la figure ci-dessus (Fig. 6), l’accommodation chez les poissons osseux (téléostéens) et chez les requins (élastomobranches) n’obéit pas au même fonctionnement : une mécanique différente a été prévue par le Constructeur.
Pour les requins et les raies (à droite) le cristallin est maintenu par deux ligaments suspenseurs reliés à deux muscles protracteurs, alors qu’il n’y a qu’un seul ligament chez les poissons osseux et un seul muscle rétracteur (à gauche).
Considérons de façon plus détaillée des caractéristiques de l’œil de l’oiseau, autre vertébré, et de ses annexes. Chezl’oiseau, à côté du schéma général qui est similaire, il est à noter des éléments propres à l’ordre et des différences entre les espèces d’oiseaux qui ne sont pas accessoires et qui nécessitent une information génétique développée de façon coordonnée pour les différents organes.
D’un point du vue anatomique, histologique et fonctionnel, l’œil des oiseaux est assez semblable à celui des mammifères. Il présente cependant des variations histologiques et fonctionnelles qui sont liées à leur mode de vie ailé. L’œil des oiseaux est, en effet, extrêmement développé́ et de grande taille. La grande taille des yeux des oiseaux nécessite une grande taille de l’orbite, ce qui entraîne un déplacement caudal de l’encéphale. Chaque espèce aviaire possède des yeux adaptés à son milieu de vie, ses mœurs, sa vitesse de déplacement, et à la recherche de la nourriture qui peut être cueillie ou chassée. Les oiseaux ont une vision précise, nécessaire pour informer leur cerveau de leur position en vol et pour donner l’emplacement des objets environnants. Contrairement à l’Homme, des oiseaux voient les couleurs dans l’ultraviolet proche.
Le vaste monde des oiseaux comprend 28 Ordres et plus de 9 000 espèces aux modes de vie multiples : terrestres (diurnes ou nocturnes) et aquatiques.
Les yeux des oiseaux sont caractérisés par leur grand volume. En effet, ils sont volumineux à la fois par rapport à la taille totale de l’animal, mais aussi par rapport à celle de la tête et donc de l’encéphale. Par exemple, le rapport entre le poids de l’œil et celui de la tête chez l’Homme est de 1 %, chez le Hibou adulte de 7 % et chez le Hibou jeune de 12 % !
Les yeux de la gent ailée sont situés latéralement ou frontalement, selon la forme de la tête. Ils sont très frontaux chez les rapaces nocturnes et plus latéraux chez les oiseaux diurnes.
Chez les rapaces, qu’ils soient diurnes ou nocturnes, l’angle que font, entre eux, les deux axes optiques est souvent voisin de 90°. Les yeux des espèces diurnes apparaissent petits pour l’observateur, en raison de la faible ouverture aux paupières, mais la partie postérieure du globe, et donc la rétine, est bien plus grande.
L’adaptation à la vie nocturne, chez les rapaces, se marque par l’accroissement du volume des yeux, à un tel point que la taille des deux yeux dépasse celle de l’encéphale. Cette adaptation à la vie nocturne est surtout caractérisée par l’importance de la longueur bulbaire axiale. Une grande longueur bulbaire axiale permet à l’œil de mobiliser un maximum de photorécepteurs, donc de recevoir l’image avec une meilleure définition, mais l’expose à des traumatismes fréquents. Inversement, les oiseaux à longueur bulbaire axiale réduite ne peuvent se guider qu’à la lumière du jour.
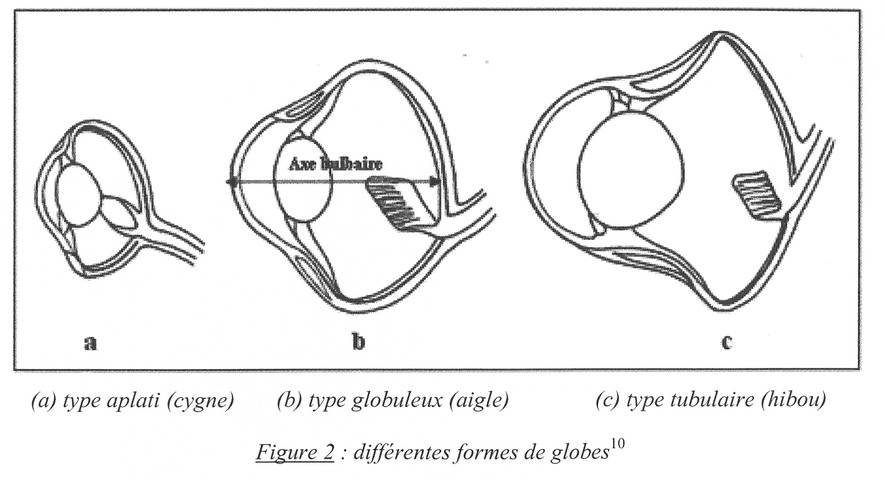
Fig. 7. La forme du globe oculaire varie selon les oiseaux : cygne (a) ; aigle (b) ; hibou (c).
On peut décrire schématiquement trois formes de bulbe oculaire chez l’oiseau :
- le type aplati (le plus fréquent, Fig 7a) : oiseaux diurnes avec une tête étroite (Colombiformes, etc.) ;
- le type globuleux ou conique : oiseaux diurnes à tête large (p. ex. Falconiformes : faucon, Fig. 7b) et à cristallin lenticulaire ;
- le type tubulaire : oiseaux nocturnes (ex. : Strigiformes : hibou, Fig. 7c) avec un cristallin sphérique et une chambre antérieure profonde.
Le bulbe lui-même a une grande rigidité. La sclère (le « blanc » de l’œil chez l’Homme) joue le rôle d’une enveloppe protectrice. Particularité anatomique des oiseaux : c’est le « squelette » de l’œil. Leur sclère est renforcée antérieurement, dans la portion du bulbe qui n’est que partiellement protégée par l’orbite, par un anneau scléral osseux formé de dix à dix-huit osselets scléraux allégés par des alvéoles remplies d’air.
Une lame cartilagineuse, qui se superpose à l’anneau scléral osseux dans son quart postérieur et vers l’intérieur du bulbe, rigidifie le reste. Les osselets de la sclère forment un cercle et s’imbriquent les uns sur les autres. Renforçant ainsi la rigidité du globe oculaire et des corps ciliaires, ils peuvent résister aux déformations du globe oculaire lors de la contraction des muscles ciliaires…
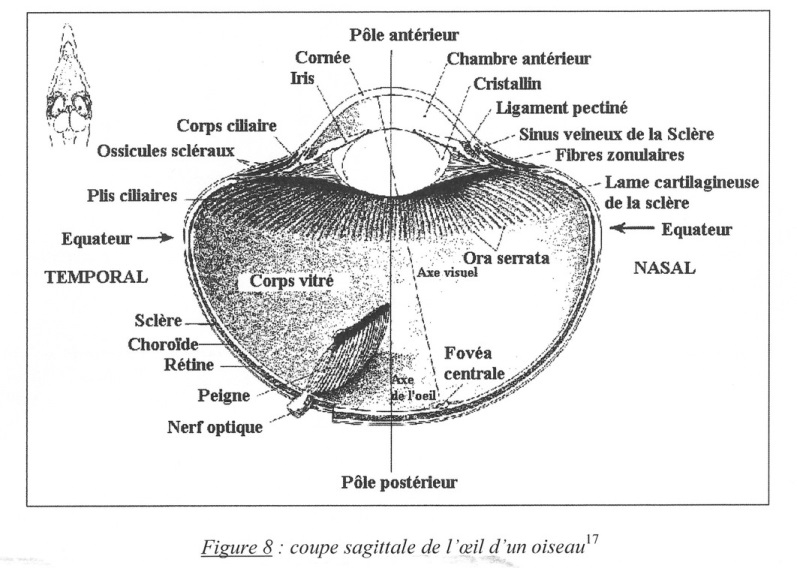
Fig. 8. Coupe sagittale de l’œil d’un oiseau.
Autre particularité de l’oiseau, lepeigne. Le peigne est un organe du fond de l’œil. Sa taille et sa forme varient d’une espèce à l’autre. Le peigne est composé exclusivement de cellules stromales pigmentées extravasculaires reliées à la rétine par des capillaires.
La taille du peigne et le nombre de plis ne dépendent pas de la taille de l’œil. En revanche, ces deux paramètres sont directement corrélés à l’acuité visuelle et à la luminosité. Les oiseaux diurnes actifs, qui ont une acuité visuelle élevée et une perception monoculaire, possèdent en général un peigne plus large et plus plissé, alors que les espèces nocturnes, qui ont une vision plus faible, ont un peigne plus petit et de morphologie plus simple.
La longueur du peigne varie entre un demi à un tiers du plus grand diamètre de l’œil. Les oiseaux chanteurs, ainsi que le Corbeau, ont le plus grand nombre de plis (30 chez ce dernier) ; les oiseaux nocturnes sont ceux qui en possèdent le moins. On dénombre 4 à 8 plis chez les Strigiformes et une vingtaine chez les Falconiformes. En général, les rapaces diurnes ont des peignes plus grands et plus plats que les rapaces nocturnes.
Avant d’évoquer d’autres différences entre oiseaux, il est intéressant de signaler un organe annexe qui existe chez d’autres animaux : la membrane nictitante. Cette petite membrane ne leur est pas accessoire et se complète de la glande de Harder, de muscles, tendons et connexions nerveuses.
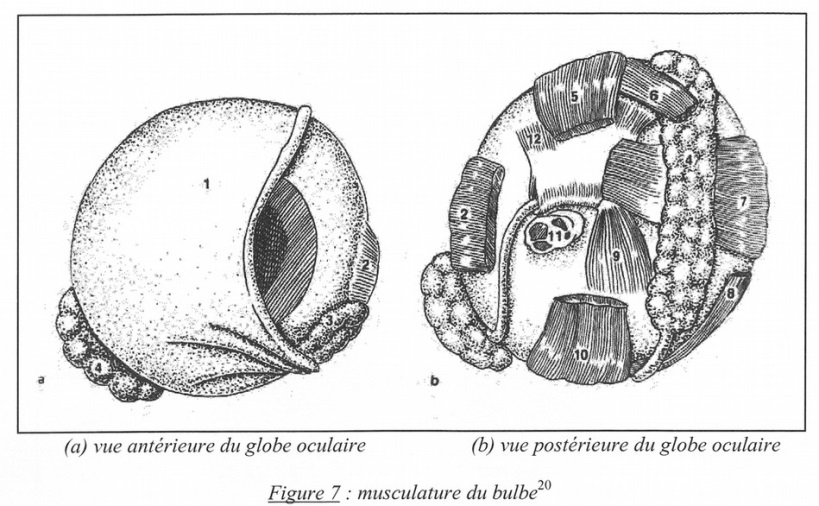
Fig. 7. Musculature du bulbe : pas si simple ! Vue antérieure (à gauche) et vue postérieure (à droite).(1)membranenictitante(2)muscledroitlatéral(3)glandelacrymale (4) glande de la membrane nictitante (5) muscle droitdorsal (6) muscle oblique dorsal (7) muscle droit médial (8) muscleobliqueventral(9)musclepyramidalis(10)muscledroitventral (11)nerfoptique(12)musclequadratus.
L’œil aviaire possède trois paupières. Les paupières supérieures et inférieures [à l’exception du Hibou et à la différence des mammifères, la paupière inférieure est toujours plus grande, plus mobile et plus transparente que la paupière supérieure. Cette dernière est, quant à elle, plus épaisse] se déplacent verticalement, et la membranenictitante(outroisièmepaupière)sedéplacehorizontalement.
Cette dernière est très mobile et se déplace très rapidement sur la cornée, à partir d’une ligne de départ dorso-nasale, et dans une direction ventro- temporale. Elle permet la protection de la cornée. Les balayages répétés répartissent les larmes sur la cornée et chassent les poussières. Les muscles de la membrane nictitante sont constitués du pyrimidalys et du quadratus, tous deux issus du pôle postérieur de l’œil. Le quadratus forme une sorte de poulie à travers laquelle le piramidalys redirige ses forces le long d’un tendon de grande taille. Ces muscles permettent de fermer la membrane nictitante sans bouger la paupière inférieure mobile. La membrane nictitante est transparente : ainsi les oiseaux peuvent-ils garder les yeux ouverts pendant le vol. Elle évite le dessèchement de la cornée puisqu’elle étale le film lacrymal en plein vol sans que l’oiseau ait besoin de fermer ses paupières. Ce rôle explique la faible fréquence des clignements palpébraux chez les oiseaux. De même, elle protège l’œil pendant le plongeon pour que l’oiseau aquatique (tel qu’un Martin-Pêcheur) puisse continuer à viser un poisson. La membrane nictitante des oiseaux marins joue le rôle d’une lentille de réfraction. Elle est plus transparente que celle des oiseaux non marins et possède en son centre une fenêtre totalement transparente.
La membrane nictitante forme une poche dans laquelle les canaux de la glande de Harder vident leurs sécrétions pour lubrifier les paupières et nettoyer la cornée. Lors de la rétraction de la membrane nictitante, les poussières et les fluides sont emmenés dans l’angle médial des paupières et passent dans le conduit lacrymo-nasal, ou bien sont expulsés dans le canthus interne.
La structure histologique de la rétine [un millimètre carré de rétine chez une Buse comporte un million de cellules visuelles comparé à 200 000 chez l’Homme] chez les diverses espèces d’oiseaux est le reflet de leur mode de vie, en particulier le mode de vie diurne ou nocturne. La rétine présente diverses organisations ou répartitions. Un exemple lié à l’acuité visuelle de l’oiseau l’illustrera : la présence d’une fovéa, de deux fovéas ou d’aréas.
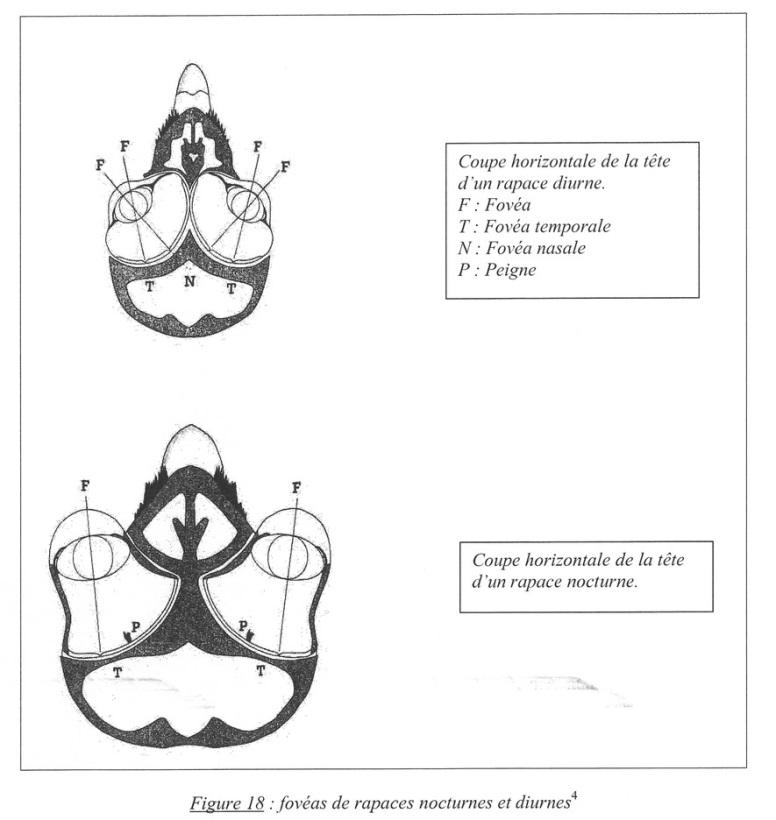
Fig. 8. Fovéas de rapaces diurnes (en haut) et nocturnes (en bas).
F : fovéa ; T : fovéa temporale ; N : fovéa nasale ; P : peigne.
La fovéa, correspond à une zone d’amincissement de la rétine où la résolution est maximale, c’est-à-dire où la vision des détails est la plus précise. Elle est située dans le prolongement de l’axe optique de l’œil. La fovéa est peuplée quasi uniquement de cônes ; et c’est dans cette zone que la majeure partie de l’appréciation des couleurs est réalisée.
Les oiseaux à vision monoculaire ont une seule fovéa centrale. Chez plusieurs oiseaux (rapaces, Hirondelles, Colibris), il existe deux fovéas dans chaque œil [Fig. 8] : l’une, déportée vers les tempes, qui sert pour la vision binoculaire ; l’autre, axiale, joue un rôle pour la vision monoculaire [beaucoup d’oiseaux voient avec un seul œil. Les mouvements oculaires des oiseaux ne sont ni synchrones ni symétriques ; un œil peut rester fixe alors que l’autre bouge. Ce sont des mouvements indépendants. Des mouvements de la tête permettent de compenser les mouvements limités du globe oculaire. Des rotations extrêmes de la tête sont constatées : 270° chez certaines chouettes !]. Ces oiseaux sont particulièrement bien équipés pour chasser en plein vol. Outre ces deux fovéas, les oiseaux de proie diurnes possèdent une « area » linéaire qui les rejoint. Il s’agit d’un épaississement local de la rétine, sans doute un dispositif améliorant l’acuité visuelle.
Les oiseaux diurnes, dont les yeux sont positionnés plus latéralement, ont une vision binoculaire restreinte à la zone située devant leur bec. Les objets situés dans cette région peuvent former des images sur les deux fovéas latérales (ou temporales) simultanément. La fovéa qui se trouve près du centre de la rétine, c’est-à-dire la fovéa centrale (ou nasale), est toujours plus dense en cônes et correspond à la vision monoculaire latérale. Sur les fovéas centrales ne peuvent se former simultanément les images d’un même objet, c’est pourquoi on appelle aussi la fovéa centrale : fovéa monoculaire. Cela explique probablement pourquoi les oiseaux penchent la tête quand ils regardent quelque chose attentivement.
Les rapaces nocturnes ne possèdent que des fovéas latérales peu différenciées, moins creusées, moins denses en cellules photo réceptrices, avec une prédominance de bâtonnets – voire, peut-être, une absence de cônes chez certaines espèces. Leur vision binoculaire est caractérisée par une acuité visuelle faible et par une sensibilité élevée.
Quelques oiseaux diurnes ne possèdent pas de fovéa dans leur rétine. Une zone spécialisée : l’area centrale fait alors office de fovéa et permet la vision du détail. Ils possèdent souvent plusieurs areas. Les oiseaux diurnes, réputés pour avoir les areas les plus perfectionnées, peuvent avoir jusqu’à trois areas distinctes.
La cornée fait partie, avec le cristallin, du système optique. Plusieurs phénomènes sont impliqués dans l’accommodation : la modification du rayon de courbure de la cornée (accommodation cornéenne), et la déformation du cristallin (accommodation lenticulaire). Selon les oiseaux, l’une ou l’autre sera privilégiée.
La cornée des oiseaux, comme celle des mammifères, est transparente, incolore et non vascularisée. Chez l’oiseau, sa courbure peut être modifiée grâce aux contractions du muscle ciliaire (muscle de Crampton) situé dans la tunique moyenne, qui tire en arrière la région du limbe. Les oiseaux nocturnes sont équipés d’une cornée de grande taille car ils ont besoin d’une grande image pour mieux voir en basse luminosité. Les oiseaux diurnes ont une cornée beaucoup plus petite. Chez beaucoup d’entre eux, la cornée a une puissance de réfraction plus grande que celle du cristallin.
Le cristallin est une lentille biconvexe, transparente, qui peut subir de légères déformations nécessaires à l’accommodation visuelle, sous l’action des muscles striés ciliaires.
Le cristallin des oiseaux, de consistance plus molle que celui des mammifères, est constitué de deux parties séparées par une vésicule : une partie centrale formée de fibres concentriques et une partie périphérique ou bourrelet annulaire, dans lequel les fibres sont plus grandes et disposées radialement. Le bourrelet, dont la présence est spécifique des oiseaux, est formé d’une seule couche de cellules hautes et constitue un anneau équatorial, peu épais chez les rapaces nocturnes et beaucoup plus épais chez les rapaces diurnes, ce qui donne la forme relativement aplatie au cristallin de ces oiseaux. Entre la masse centrale du cristallin et le bourrelet annulaire se trouve la vésicule du cristallin, considérée naguère comme un artéfact, remplie d’un liquide de nature protéique. Elle contribue à la mollesse d’ensemble du cristallin et par là, semble-t-il, à la rapidité de l’accommodation.
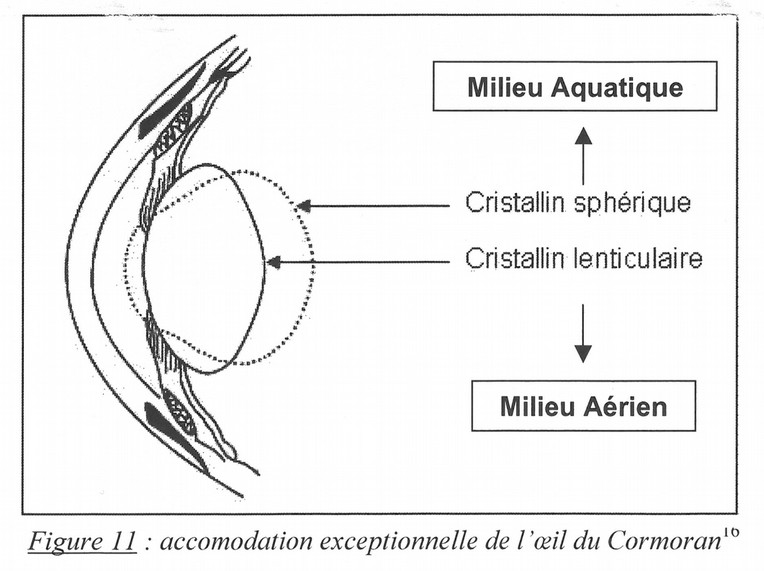
Fig. 9. Accomodation spécifique à l’œil du Cormoran, selon le mileu où il opère.
Chez les oiseaux plongeurs, le sphincter irien est si puissant que l’iris peut déformer le cristallin, alors très mou, en se contractant. L’iris recouvre les marges du cristallin et sa contraction-dilatation fait varier le diamètre et la forme de la pupille. En effet, les oiseaux marins transforment leur cristallin lenticulaire (de la vision aérienne, pour viser et plonger) en cristallin sphérique (de la vision aquatique, pour nager et chasser sous l’eau)](Fig. 9).
La musculature de l’iris est striée et soumise à la volonté chez les oiseaux.
Signalons aussi que :
- certains oiseaux ont une glande à sel (ou glande nasale). Cette glande est présente chez les oiseaux marins et chez certains oiseaux comme la Perruche ondulée. Elle leur permet d’éliminer le sel absorbé après consommation d’une eau salée ;
- les gouttelettes lipidiques près des cônes permettent, avec des pigments, une vision pentachromatique pour certains [sensibilités aux UVA (320 à 400 nm)] et favorisent le contraste des images ;
- la position des yeux frontaux ou latéraux corrélée avec la vision binoculaire et la vision monoculaire (panoramique) est essentielle pour la survie ;
- les champs visuels sont liés à la forme des yeux et à la position de ceux-ci dans le crâne ;
- la décussation des fibres nerveuses au niveau du chiasma du cerveau est, selon les oiseaux, partielle ou totale…
Les yeux composés des arthropodes (crustacés, arachnides, insectes…) : apposition ou superposition
L‘œil composé d’un arthropode est formé par la juxtaposition d’unités photoréceptrices, les ommatidies, constituées chacune comme un œil élémentaire : une cornée faite de cuticule transparente, un cristallin transparent et conique, 8 cellules rétiniennes. Ces dernières possèdent des pigments photorécepteurs dans des microvillosités membranaires qui, en s’emboîtant les unes dans les autres, forment un bâtonnet photorécepteur. Le cristallin ne se déforme pas : ici, il n’y a pas d’accommodation, mais la convergence des lentilles que forment cornée et cristallin, et la longueur du bâtonnet photorécepteur permettent de recevoir sur celui-ci les rayons lumineux émis à différentes distances.
Comme chez les vertébrés, un stimulus lumineux, à la suite d’une réaction photochimique du pigment photorécepteur, génère un message nerveux qui, propagé jusqu’aux centres nerveux, est perçu comme une image élémentaire. Il existe en outre des pigments non photorécepteurs formant un écran autour de chaque ommatidie. Leur disposition varie en fonction de la luminosité du milieu de vie de l’animal.
La Fig. 10 illustre le fait que, bien qu’organisés suivant le même modèle, les yeux du Criquet et ceux de l’Écrevisse fonctionnent différemment.
Chez le Criquet, les pigments écrans dispersés isolent chaque ommatidie de ses voisines. La lumière en provenance d’un point du champ visuel traverse la cornée et le cristallin d’une ommatidie et ne vient stimuler que le bâtonnet photorécepteur de cette ommatidie.
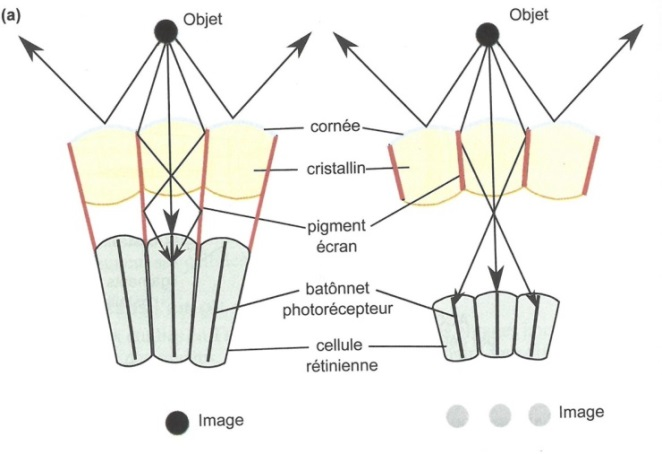
Fig. 10. Les yeux composés des arthropodes. Ils fonctionnent :
-soit par apposition, en milieu terrestre éclairé (le Criquet, à gauche) ;
-soit par superposition, en milieu aquatique peu lumineux (l’Écrevisse, à droite).
Le traitement par les centres du message nerveux correspondant permet alors de percevoir ce point. Il existe des milliers d’ommatidies et les centres visuels traitent l’ensemble des messages nerveux qu’elles envoient pour reconstituer une image formée d’une mosaïque de points lumineux juxtaposés. L’œil du Criquet fonctionne ainsi par apposition, ce qui permet la formation d’images précises mais peu lumineuses, puisqu’une partie des rayons émis par les objets est absorbée par les pigments écrans. Chez l’Écrevisse, les bâtonnets photorécepteurs sont courts et les pigments écrans sont concentrés autour du cristallin, ce qui permet aux rayons lumineux qui ont traversé une ommatidie d’aller aussi exciter les bâtonnets photorécepteurs des ommatidies voisines. L’image perçue est donc moins nette que pour le Criquet puisqu’un même point lumineux peut ainsi être perçu comme plusieurs points. Dans ce fonctionnement par superposition, 50 % de la lumière qui stimule un bâtonnet photorécepteur a traversé d’autres ommatidies que la sienne.
La convergence, sur un même bâtonnet photorécepteur, de rayons lumineux issus de différents points du champ visuel rend plus probable la photoréception dans un milieu peu lumineux. Lorsque l’Écrevisse est dans un milieu bien éclairé, les pigments écrans se dispersent, permettant un fonctionnement par apposition. À l’inverse, le fonctionnement par superposition existe aussi chez les insectes terrestres vivant la nuit ou dans un milieu peu lumineux.
Notons ici que l’œil de la Langouste [cf. Le Cep n°11] (et des animaux voisins) ainsi que celui de la Coquille Saint-Jacques sont parmi les plus remarquables que l’on puisse trouver dans l’ensemble de la biosphère ! L’un des traits les plus frappants de l’œil de la Langouste, que l’on perçoit immédiatement même à l’observation superficielle, est que ses facettes constituent descarrésparfaits. La rareté de cette forme en biologie fait que cet œil ressemble plus à un appareil artificiel qu’à une structure naturelle.
En résumé…
Il existe plusieurs types d’yeux chez les animaux (œil simple ou composé). Dans chaque type, le fonctionnement de base est le même, mais la structure (paupières, rayon de courbure de la cornée, mouvements du cristallin, disposition des pigments, etc.) est pré-adapté pour convenir au milieu de vie de l’animal. À cause des différences d’indice de réfraction des milieux, un œil de vertébré aquatique ne peut pas voir nettement les objets terrestres – et réciproquement – s’il n’a pas des annexes le lui permettant (membrane nictitante, cristallin qui devient lenticulaire grâce aux muscles présents, etc.), point de fixation des muscles avec des tendons qui fonctionnent quelquefois comme des poulies, voies nerveuses, etc. Tout est pré-adapté pour la survie de l’individu à partir d’une cellule-œuf organisée et d’un programme génétique qui va être lu de façon différentielle selon les régions de l’organisme.
Les différences inter-spécifiques ne laissent guère imaginer que l’évolution graduelle des êtres fût possible par le simple fait de mutations aléatoires qui, ponctuelles et locales (les versions alléliques en sont des exemples), ne changent pas grand-chose à l’échelle de l’organisme entier, quand même elles ne lui sont pas funestes. C’est, à chaque fois, toute une réécriture du message contenu dans l’ADN au sein d’une cellule déjà très spécifique, l’ovule puis la cellule-œuf, que le transformisme doit reconsidérer.
« Quam magnificata sunt opera tua Domine : nimis profundæ factæ sunt cogitationes tuæ ! Vir insipiens noncognoscet, et stultus non intelliget hæc » [Que vos œuvres sont grandes Seigneur, que vos pensées sont profondes ! L’homme sans sagesse n’y connaît rien, et l’insensé n’y peut rien comprendre] (Ps 91, 6-7).
Bibliographie
- BCPST 1e année, Dunod, Chapitre 12, p. 364-366.
- NOIRE Nathalie, Ophtalmologie des oiseaux de compagnies et des oiseaux sauvages, thèse pour le diplôme d’Etat de Docteur vétérinaire présentée le 21/01/2008 devant la Faculté de Médecine de Nantes. http://www.sferov.org/pdf/These_Nathalie_Noire.pdf
1 Del’Univers à Dieu, Montsûrs, Résiac, 1978. Cf.« L’œil et la vision », LeCep n°9, novembre 1999, p. 80.
2 Merveillesde l’œil, Paris, Bloud, 1911. Cf. « L’œil est fait pour voir », Le Cep n°47, 48 et 49, avril à novembre 2009.
3 Cf. W. GITT, « Le cristallin : génial ! », Le Cep n°45, octobre 2008, p. 81.
4 Cf. G. SALET & L. LAFONT, « La collaboration de la science et de la théologie », Le Cep n°20, août 2002, p. 15.