Partager la publication "Retour sur la réalité des espèces"
Par : Dominique Tassot
Présentation : Depuis Lamarck et Darwin, l’idée que les espèces vivantes sont des formes transitoires et que la vie est un continuum s’est si bien imposée que plus personne n’en mesure l’incongruité. Un article récent (mai 2018), publié dans la revue Human Evolution, présente les travaux faits depuis 15 ans sur un petit gène mitochondrial, la cytochrome oxidase de type 1 (COI), par deux chercheurs, l’un américain, Mark Stoeckle (université Rockefeller), l’autre suisse, David Thaler (université de Bâle). Le COI permet une identification fine des espèces, ce qui confirme la réalité objective des classifications taxinomiques. Surtout, l’étude constate une claire séparation des espèces : même entre espèces les plus voisines, les écarts des séquences sont de l’ordre de 4 % à 10 %, tandis que les variations internes à l’espèce sont minimes (0,2 % en moyenne). Les espèces sont comme « des îles [bien distinctes] au milieu de l’espace ». Pour sauver le mythe évolutionniste, les auteurs ont imaginé que la divergence étudiée aujourd’hui n’aurait duré que très peu de temps (à l’échelle géologique). Il est piquant de voir lancer cette hypothèse par des gens appartenant à une « lignée » de chercheurs, lesquels, depuis deux siècles, fondent la crédibilité de leur thèse sur des durées si longues que l’imagination s’y perd !
Le regretté Pierre-Paul Grassé (1895-1985), pourtant titulaire à la Sorbonne de la chaire d’évolution des êtres organisés, était connu pour dire : « Si l’on voulait démontrer la stabilité des espèces, on ne trouverait pas meilleur argument que toutes les expériences de mutations artificielles1 faites depuis 50 ans. » En effet, les malheureuses drosophiles et les colibacilles mutés, quand ils ont pu se reproduire, n’ont jamais donné naissance qu’à des colibacilles et à des drosophiles. Grassé pourrait aujourd’hui y ajouter un article étonnant, paru en mai dernier dans Human Evolution sous le titre : « Pourquoi les mitochondries devraient-elles définir les espèces ?
Cette fois, en effet, il ne s’agit plus seulement de la stabilité des espèces, mais de leur réalité ontologique, même si les auteurs signalent ce point mais ne s’y attardent pas ; il sera aisé de comprendre pourquoi.
La revue ayant accepté la publication s’intitule Human Evolution. Il s’agit donc d’expliquer les faits, mais selon la grille d’interprétation de l’évolutionnisme. Or les faits ne « collent » pas avec la théorie2. La presse n’a d’ailleurs retenu que deux affirmations spectaculaires. La première est que « 90 % des animaux, ou plus exactement 9 espèces animales sur 10, sont apparus sur Terre à peu près en même temps que l’homme, il y a 100 000 ou 200 000 années. L’un des deux auteurs, David Thaler, généticien à l’université de Bâle, reconnaît que cette conclusion de sa recherche est “ très surprenante ” : “ Je l’ai combattue autant que je l’ai pu ”, avoue-t-il dans un entretien accordé à l’AFP »3. La seconde est que « pour les 7,6 milliards d’hommes de la planète, les 500 millions de moineaux et les 100 000 bécasseaux, la diversité génétique “est presque identique” a déclaré Mark Stoeckle, de l’université Rockefeller de New-York, à l’AFP4 ». Mais, avant de commenter ces affirmations, il convient d’établir le sérieux de l’article.
Les deux chercheurs en question sont des universitaires reconnus ; ils avaient publié ensemble en 2014 un article soulevant déjà le lièvre : Le codage à barres de l‘ADN [séquençage] marche en pratique, mais pas selon la théorie [des mutations neutres]4. Mark Stoeckle, l’auteur américain, travaille sur le sujet depuis longtemps, puisqu’il donnait en 2003 à la revue BioScience un article intitulé « La Taxonomie, l’ADN et le code-barre de la vie5 ». On ne peut d’ailleurs soupçonner l’université Rockefeller où il travaille de vouloir dénigrer l’évolutionnisme : elle fut fondée en 1901 par John D. Rockefeller, et rappelons que les fouilles de Chou-Kou-Tien (« l’Homme de Pékin ») furent financées par la Fondation Rockefeller6.
Les auteurs s’efforcent d’ailleurs, tout au long de l’article7 et avec plus ou moins de succès, de montrer qu’une interprétation évolutionniste demeure possible. Le travail accumulé est colossal. Il l’est en extension : 162 articles, la plupart récents, sont cités et mis en références. Il l’est aussi en compréhension : on trouve nombre des classiques de la biologie, car la question de fond – le statut de « l’espèce » – traverse inévitablement la pensée des grands noms de cette science. Comparaissent ainsi Mayr (1955), Bateson (Cambridge, 1908), Gould (2002), Dobzhansky (Columbia, 1937), Lewontin (1991), Kimura (1989), Crick (1966), John Maynard Smith (1974), etc. Une référence à Thomas Kuhn8 manifeste une nette conscience de l’enjeu : « Dans La Structure des Révolutions scientifiques, Thomas Kuhn montre comment chaque modèle scientifique prend certains faits naturels ou certains résultats expérimentaux comme les données-clés à expliquer. Nous considérons la structure en amas séparés [cluster] des séquences de la cytochrome oxydase – faible variance au sein des espèces et souvent, mais pas toujours, fossé avec les espèces les plus proches – comme un fait fondamental qu’un modèle de l’évolution ou de la spéciation doit expliquer » (ST, p. 8 et 10). En clair – disent les auteurs –, nous sommes devant un « fait crucial » de nature à remettre en cause le modèle évolutionniste, si celui-ci n’en donnait pas un jour une explication satisfaisante.
De quoi s’agit-il, au fond ? Rien de moins que de la réalité des espèces. Darwin écrivait à Asa Gray9 le 21 décembre 1859, donc aussitôt après la publication de son De l’Origine des espèces : « Je pense qu’il est très important que mes idées soient lues par des hommes intelligents, accoutumés aux arguments scientifiques, tout en n’étant pas10 naturalistes. Cela peut paraître absurde, mais je m’imagine que de pareils hommes entraîneront après eux les naturalistes qui s’entêtent à croire qu’une espèce est une entité11.»
Depuis un siècle, avec le rouleau-compresseur darwinien, la question pouvait sembler tranchée et donner la victoire au nominalisme12 : seuls existeraient réellement les individus, multiples êtres en transit entre une forme ancestrale inconnue et une forme à venir encore indéterminée, mais que, par nécessité pratique, nous regroupons selon leur apparence (le « phénotype »). Ces groupes artificiels, qualifiés par nous d’espèces, ne seraient donc pas des réalités substantielles (des « entités » », écrit Darwin), mais des catégories abstraites, créées pour notre commodité intellectuelle. Ce nominalisme biologique semblait d’autant plus certain que les taxonomistes sont partagés sur la manière de définir l’espèce, la définition la plus connue, attribuée à Mayr, consistant en l’interfécondité entre individus de la même espèce. Mais on compte une trentaine de définitions différentes de l’espèce (ST p. 8) et l’idée que les espèces sont transitoires a beaucoup fait pour décourager les vocations de taxonomistes, d’autant qu’il s’agit d’une science ardue exigeant un grand effort de mémoire. Il est notable cependant que, dans la pratique, l’étude des espèces n’a jamais été détrônée : indispensable, elle avait même connu un renouvellement avec l’apparition de la biologie moléculaire dans les années 1970.
L’interfécondité implique la filiation et donc la transmission du génome. Sans prétendre réduire l’être vivant à ses chromosomes, il s’avérait que l’ADN portait bel et bien des caractères pertinents reliés aux principaux traits d’un individu. L’analyse de l’ADN s’immisça donc dans nombre de questions pratiques, comme la détermination de l’hérédité, preuve aujourd’hui admise en Justice.
Mais le génome complet était long et coûteux à déchiffrer (à séquencer). On découvrit alors qu’une protéine, le cytochrome C, ne comportant qu’une centaine d’acides aminés et facile à localiser, se rencontrait chez presque tous les organismes, depuis les bactéries jusqu’aux mammifères. Or il est facile de comparer deux cytochromes : il suffit de compter les bases différentes entre leurs deux séquences mises en parallèle et de diviser le résultat par le nombre total de bases. Le ratio obtenu mesure la divergence. On dispose ainsi d’un critère simple et quantifié, ce qui ne peut que stimuler la recherche. Les divergences mesurées sont minimes au sein de l’espèce et significatives entre deux espèces même voisines.
En 1972, Margaret O. Dayhoff publiait une matrice à 1 089 entrées donnant les divergences (en %) des séquences sur 33 cytochromes appartenant à des espèces très diverses : mammifères, poissons, insectes, plantes, levures et bactéries13. Sans surprise, on constate que les espèces peuvent se regrouper selon les classements traditionnels : « par exemple, entre un cheval et un chien (deux mammifères), la divergence est de 6 % ; entre un cheval et une tortue (deux vertébrés), de 11 % ; entre un cheval et une mouche (deux animaux), de 22 % » (ETC, p. 287-8).
Si donc « l’arbre généalogique » de la vie correspondait à une réalité, cette corrélation entre le taux de divergence et la proximité taxonomique devrait se retrouver dans l’ensemble de l’arbre : on devrait trouver moins de divergences entre les êtres plus « simples », et donc plus « primitifs » que sont les bactéries et les levures, qu’entre une bactérie et un sapin ou, a fortiori, entre une bactérie et un pigeon. Or il n’en est rien : tous les organismes ayant un noyau cellulaire (eucaryotes) sont à égale distance de la bactérie Rhodospirillum rubrum (ETC, p. 289).
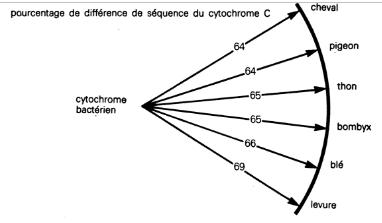
Selon Denton, « ce résultat doit être tenu pour une découverte des plus étonnantes de la science moderne… Le règne bactérien n’a aucun voisin parmi la fantastique diversité des types eucaryotes. Les “ chaînons manquants ” sont bel et bien manquants » (ETC, p. 289).
Et le même phénomène se reproduit à un niveau taxonomique moindre, ainsi entre le bombyx et divers vertébrés :
Il en résulte un modèle de la nature clairement conforme à la typologie classique fondée sur les différences morphologiques, sans le moindre indice d’une continuité biologique entre les classes et les autres groupes.
La série évolutive bien connue est la suivante :
cyclostome > poisson > amphibien > reptile > mammifère.
Or l’Atlas de Dayhoff, publié en 1978, donne les divergences suivantes (ETC, p. 292) :
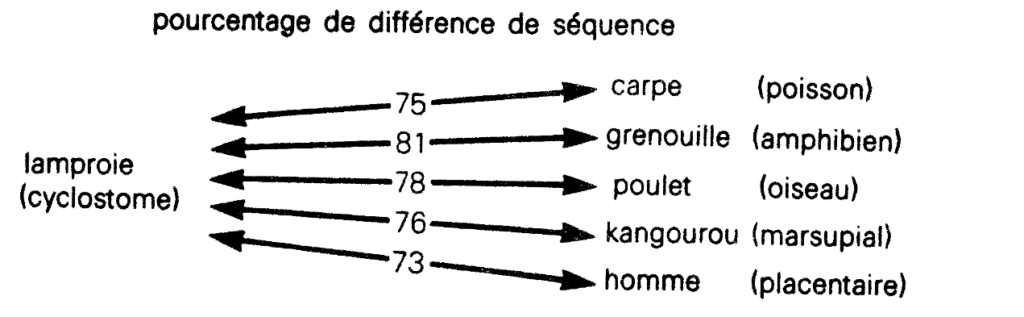
Nous ne retrouvons donc nullement l’écart croissant attendu. De même les vertébrés terrestres apparaissent tous équidistants du poisson (ETC, p. 293) :

À l’époque, cette contradiction flagrante avec la prédiction évolutionniste fut passée sous silence. En 1969, Margaret O. Dayhoff affirma placidement qu’« un arbre généalogique construit de cette manière avait la même typologie que celle déduite des considérations morphologiques conventionnelles14 ». Ce n’était pas entièrement faux puisque, nous l’avons vu, les êtres vivants se classaient en groupes voisins correspondant à la taxonomie ; mais on n’y trouvait pas le moindre indice « généalogique ».
Le fameux « arbre de la vie » n’a d’ailleurs jamais comporté que des rameaux terminaux ; les branches et le tronc sont de pures abstractions, comme nombre de biologistes en avaient depuis longtemps fait la remarque.
Dans ce contexte, quelles informations nouvelles vient donc apporter cet article de Stoeckle et Thaler ? Il permet tout d’abord d’actualiser et d’affiner les considérations antidarwiniennes faites il y a 30 ans par Sermonti et Denton. La présente étude porte cette fois sur un autre petit gène, la cytochrome oxydase de type 1 (en abrégé COI ou MT-CO1)15. Il a été séquencé couramment à travers le monde, chez de multiples individus appartenant à de nombreuses espèces, si bien que la matière disponible est considérable : 5 millions d’ADN divers ont été analysés, répartis dans la majorité des phylums animaux, de sorte que l’échantillon représente valablement environ les trois quarts des espèces connues (ST p. 7).
L’accord entre les regroupements faits en considérant les séquences moléculaires et les groupes taxonomiques de la zoologie est amplement confirmé, y compris par les exceptions : ce sont là précisément où les spécialistes sont en désaccord sur la définition de l’espèce, ou encore sur les groupes mal étudiés que les écarts augmentent.
De plus, la quantité des données disponibles permet le calcul de moyennes statistiques : on chiffre ainsi avec précision l’ampleur des différences tant au sein de l’espèce qu’entre les espèces voisines. Les variations intraspécifiques sont minimes (en moyenne, entre 0,0 % et 0,5 % ; 0,1 % chez l’homme, le plus étudié) (ST p. 10). Ci-après l’histogramme des écarts moyens (notés APD : average pairwise difference) mesurés par les paires reconnues différentes tout en occupant la même place dans la séquence de 648 paires (ST p. 4).
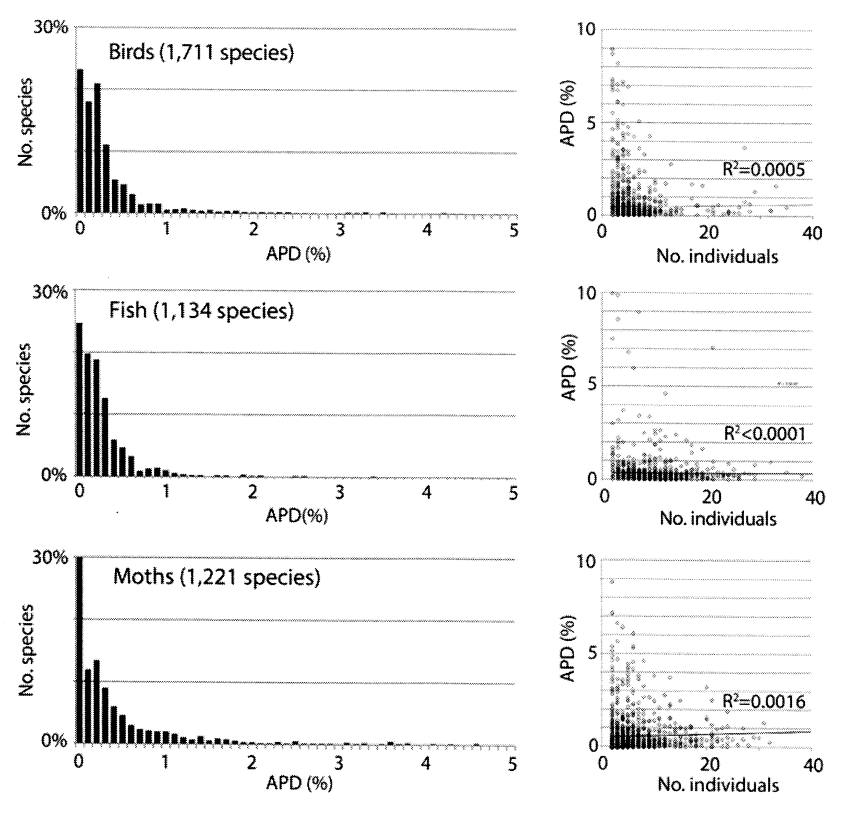
« Fig. 1. Une faible variabilité du COI est la norme chez les animaux et ne peut résulter ni d’un biais dans le choix des exemples ni d’échantillons trop faibles » (ST p. 4)
On notera (colonne de droite, APD en ordonnées) que peu d’individus dépassent 1 % d’écart : les moyennes sont donc significatives. De plus (colonne de gauche), le nombre d’espèces étudiées (1 711 pour les oiseaux, 1 134 pour les poissons et 1 221 pour les papillons de nuit) permet des conclusions d’une grande généralité : les écarts moyens supérieurs à 1 % (APD en abcisse) sont minimes.
La figure suivante compare cette fois les écarts intraspécifiques (W : within) et les écarts mesurés avec les espèces voisines (B : between) sur « l’arbre de la vie » (ST p.5).
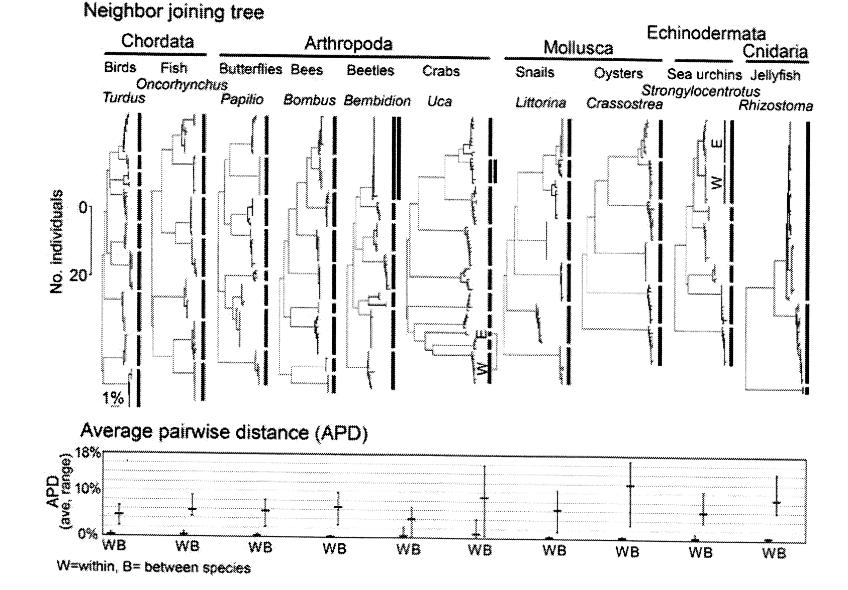
« Fig. 2 : D’amples écarts entre les espèces, si on les compare aux écarts intraspécifiques uniformément faibles, sont la norme chez les animaux » (ST p. 5).
Dans le tableau inférieur, les écarts moyens (trait gras horizontal) et la dispersion (trait fin vertical) font voir une dispersion intraspécifique (W) minime (moins de 1 %), sauf chez les scarabées (2 %) et les crabes (4 %), tandis que les écarts interspécifiques moyens (B) varient de 4 % à 6 % en général (mais 9,5 % pour les crabes et 14 % pour les huîtres). Selon l’image évocatrice proposée par les auteurs : « Les espèces sont des îlots dans l’espace des séquences » (ST p. 9).
Bref, il n’existe aucun recouvrement entre les espèces dites « voisines », et c’est précisément pour cette raison que l’analyse séquentielle du COI est si développée : elle sert à identifier une substance animale, par exemple à contrôler si un pâté d’alouette ne recèlerait pas quelques traces d’une viande de mouton.
Il suffit de s’assurer que les séquences COI présentes dans l’échantillon sont bel et bien celles spécifiques aux alouettes.
Cette utilité pratique explique la diffusion de la méthode ; elle confirme aussi sa pertinence : les organismes vérificateurs l’auraient abandonnée si elle n’était pas fiable, et il est très facile de faire un test de contrôle pour s’assurer du sérieux du laboratoire. Nous ne sommes donc pas ici dans le domaine d’une recherche fondamentale théorique et pointue, où peu de spécialistes seraient en mesure de juger d’une publication ; nous sommes devant les résultats d’une technique largement développée et consensuelle dont la conclusion paraît claire : les espèces sont une réalité connaissable et reconnaissable ; elles sont bien distinctes et valident globalement la classification de Linné et de ses successeurs. Stoeckle et Thaler en ont une claire conscience et se couvrent d’un auteur influent (publié par l’Académie américaine des Sciences en 1997) pour introduire le débat : « Dans un document fondateur de la phylogéographie, Avise et ses collègues ont noté la division séculaire de la biologie entre les lignées intellectuelles de Linné pour lesquelles les espèces sont des entités bien définies et celles de Darwin qui insistent sur les changement cumulatifs au sein des espèces, conduisant ainsi à de nouvelles espèces. Avec préscience [l’article en référence datait de 1987], ils avaient suggéré que l’analyse de l’ADN mitochondrial offrirait un pont pour franchir ce fossé intellectuel. Le séquençage [barcoding moléculaire] de l’ADN [mitochondrial] offre désormais une base de données des plus étendues permettant la concrétisation quantifiée et à l’échelle du règne animal, d’une telle vision » (ST p. 7).
Mais comment concilier vraiment ces deux écoles présentées dans l’histoire des sciences comme contradictoires : les fixistes (Linné)16 et les évolutionnistes (Darwin) ? Nos auteurs vont le tenter, indirectement, par la démographie historique : puisque la dérive génétique chez l’homme moderne est de 1 % par million d’années [étrangement cette affirmation est donnée sans citer une source en référence], et que l’écart intraspécifique actuel est de 0,1 %, c’est que notre humanité descend d’un unique ADN mitochodrial, il y a 100 000 ans (ST p. 22). C’est la thèse du « goulot d’étranglement » : bien sûr l’humanité est plus vieille, la divergence d’avec son ancêtre animal est bien plus ancienne, cependant tous les rameaux issus de la souche primitive se seront éteints, sauf un, et c’est à partir de cette « mère unique » (ST p. 19) [le mot « Ève » n’est pas prononcé, mais on ne peut s’empêcher d’y penser !] que l’évolution aboutissant à l’humanité actuelle a repris17… Cette « solution » soulève nombre de questions irrésolues. L’une d’elles est que les espèces peu répandues ou se reproduisant moins vite devraient manifester moins de divergences que les espèces nombreuses, ce qui n’est clairement pas le cas. Comment se fait-il que les écarts intraspécifiques soient aussi voisins (inférieurs à 2 %) ? Comment chaque espèce aurait-elle connu son propre « goulot d’étranglement », mais judicieusement placé pour faire converger les écarts actuels ? Nos auteurs se contentent ici d’extrapoler aux autres espèces l’hypothèse faite pour l’homme en suggérant qu’un « âge glaciaire » eut cet effet dévastateur général (ST p. 23).
On comprend au demeurant que Stoeckle et Thaler restent ici évasifs. Comment pourraient-ils conclure et tirer les inévitables implications de leur travail sans chatouiller le dogme évolutionniste ? On sera peut-être tenté de récuser la cytochrome oxydase en notant que ce COI ne constitue jamais que 5 % du génome mitochondrial et moins de 1 % du génome total !
Toutefois nul ne prétend que le COI définisse et décrive chaque espèce : il ne fait que l’identifier, la distinguer des autres. Ce n’est donc qu’un marqueur, comme le sont nos empreintes digitales. On ne sait rien d’une personne si l’on n’en connaît que les empreintes. Mais on sait s’il est présent (s’il a touché l’arme du crime, par exemple). Dire que le COI a permis d’identifier une espèce implique l’existence réelle de cette espèce. C’est donc contredire le présupposé darwinien d’une divergence évolutive aboutissant à de nouvelles espèces par adaptations cumulatives à l’environnement.
Or il existe une solution toute simple et cohérente avec les faits observés : si les variations au sein de l’espèce demeurent dans de si étroites limites, c’est qu’il existe une « force de rappel », tel ce processus qui, après une blessure au doigt, reproduit les mêmes dessins sur l’épiderme, processus qui, ici, exclut les mutations dites « positives » (constituant un progrès) lesquelles aboutiraient à une divergence cumulative. Cette force, ne serait-ce pas, tout simplement, le primat de la forme, la réalité et donc la persistance des formes !
Stoeckle et Thaler sont prisonniers d’un monde en devenir perpétuel, d’un monde où rien n’est tout simplement ce qu’il est, d’un monde où tout procède d’autre chose (en l’occurrence d’un hypothétique ancêtre qui aurait été différent) : le monde de l’évolution.
Expliquer l’être par le devenir : tâche impossible ! Devant la perplexité de nos auteurs, concluons avec Jeanne Smits : « La réponse a toutes les chances de relever d’un autre domaine, non accessible à la science18 : quelque chose qui s’appellerait la création… »
Expliquer l’être par le devenir : tâche impossible ! Devant la perplexité de nos auteurs, concluons avec Jeanne Smits : « La réponse a toutes les chances de relever d’un autre domaine, non accessible à la science18 : quelque chose qui s’appellerait la création… »
Notes :
1- Allusions aux expériences de Morgan sur la mouche du vinaigre (Drosophile) et à celles de Lanski sur le colibacille (Escherichia coli).
2 – On devrait plutôt parler de « mythe » que de « théorie »
3 – Cf. Jeanne SMITS, https://reinformation.tv/90-especes-animales-apparues-meme-temps-evolution-etude-genetique-smits-84927-2/
4 – STOECKLE, M. Y. & D. S. THALER, « DNA barcoding works in practice but not in (neutral) theory », PLOSONE, 2014, 9(7), e 10 0755.
5 – « Taxonomy, DNA and the bar code of life », BioScience, 2003. 53 : p. 2-3.
6 – Fort opportunément, les fossiles du Sinanthrope ont disparu durant la guerre sino-japonaise, si bien qu’une mésaventure semblable à celle de l’Homme de Piltdown (reconnu comme une forgerie) lui sera épargnée : on ne peut pratiquer d’analyses physico-chimiques sur des moulages
7 – STOECKLE, M. Y. & THALER, D. S, « Why should mitochondria define species ? », Human Evolution, vol. 33-n. 1-2 (1-30)-2018, ci-après référencé dans le corps du texte par ST.
8 – KUHN T., The Structure of scientific Revolutions, Chicago U.P., 1962, p. 126.
9 – Asa GRAY (1810-1880) : botaniste américain, alors enseignant à Harvard, ayant fourni à Darwin de nombreuses observations.
10 – Souligné dans le texte.
11 – La Vie et la Correspondance de Charles Darwin (trad. Henry de Varigny), Paris, Reinwald, 1888, t. II, p. 87 ; souligné par nous.
12 – Thèse de la philosophie médiévale tardive selon laquelle les catégories de la pensée (les « universaux » en particulier) ne sont que des noms, des appellations humaines et non des réalités ontologiques. S’oppose en particulier à la philosophie dite « réaliste ».
13 – Les éléments ici rassemblés sur le cytochrome C proviennent presque intégralement de l’ouvrage de Michael DENTON : L’Évolution, une théorie en crise (1985), Paris, Londreys, 1988, ch. 12, p. 282-296, désigné par ETC dans le corps du texte.
14 – Giuseppe SERMONTI, Dimenticare Darwin, Milan, Rusconi, 1999 (trad. angl. citée : Why a Fly is not a Horse, Seattle, Discovery Institute Press, 2005, p. 154.
15 – La séquence d’ADN étudiée (5 % du génome mitochondrial) comporte 648 paires de bases (bp) seulement, ce qui permet, avec les outils actuels, un décodage rapide et bon marché.
16 – En vérité le « fixisme » caricatural d’espèces immuables n’a jamais été soutenu par personne, y compris par Linné : les êtres vivants varient (c’est là leur différence d’avec le minéral). Ce qui est en cause ici, c’est l’ampleur des variations. Linné, comme ses successeurs, distinguent les caractères principaux (quadrupède, poilu, etc.), immuables, d’avec les caractères secondaires (couleur, taille, etc.), qui eux sont mutables.
17 – Pour l’évolutionnisme, la divergence des formes vivantes est chose naturelle. Pendant deux siècles, la stabilité des espèces a donc été interprétée en disant : « Certes, à notre échelle d’observation, on ne voit pas cette divergence, mais le lent processus s’est étalé sur de si longues durées … Il est donc normal qu’il nous échappe ! » Voici aujourd’hui le discours inverse : « On ne constate pas encore la divergence, car elle ne dure que depuis trop peu de temps ! » L’esprit scientifique voudrait qu’un phénomène inobservé ne soit pas même pris en compte. Mesurons ici à quel point René Thom avait raison lorsqu’il qualifiait le darwinisme de théorie « langagière ». Ce n’est vraiment qu’un enchaînement de sophismes, mais de sophismes savants !
18 – Ndlr. Plus exactement : inaccessible à une science qui s’interdit a priori de considérer cette perspective, fût-elle – comme c’est le cas ici – l’explication la plus simple et la plus cohérente avec les faits connus.