Partager la publication "Coaptation et finalité"
Par Lucien Cuénot
Coaptation et finalité1
Lucien Cuénot2
« Les rationalistes fuient le mystère pour se précipiter dans l’incohérence » (Bossuet)
Résumé : la « coaptation », chez les êtres vivants est, pour Cuénot, l’existence d’un organe approprié à une fonction. Ainsi pelage ou plumage (organes) se retrouvent chez tous les oiseaux et mammifères, qui sont précisément les animaux homéothermes, ceux qui doivent garder une température interne constante. Il est difficile d’y voir un effet du hasard, puisque tout suggère un plan préexistant à la création de l’organe (Cuénot, étant évolutionniste, dit « apparition » de l’organe). Un exemple remarquable d’une telle coaptation est celui des callosités du Phacochère. En effet, elles sont présentes dès le jeune embryon, lequel n’a pas encore pu fouiller le sol et ainsi endurcir son épiderme aux points de frottement. La callosité est donc préparée (comme la plante des pieds chez le fœtus humain dès le 4ème mois), et il est difficile de nier ici une finalité à l’œuvre chez l’être vivant.
Pelage et plumage :
J’entends par coaptation anatomo-physiologique l’existence chez un être d’un dispositif organique à rôle passif ou actif, dont la présence rend possible un fonctionnement physiologique ou permet un certain comportement.
Un exemple sera plus clair qu’une définition compliquée : Oiseau et Mammifère dérivent assurément de Reptiles couverts d’écailles épidermiques, car on retrouve encore ces écailles reptiliennes sur les pattes des Oiseaux, et çà et là chez les Mammifères à queue dépourvue de longs poils.
Très probablement, la plume primitive résulte d’une transformation de l’écaille dont la partie distale s’est divisée en franges ; quant au poil, c’est plutôt une néoformation indépendante; ces phanères, d’origine épidermique comme les écailles, mais de structure beaucoup plus compliquée, sont actuellement des caractéristiques absolues des deux groupes supérieurs de Vertébrés. Or, Mammifères et Oiseaux, seuls dans le règne animal, sont des homéothermes gardant constante une température interne, de 29° (Monotrèmes) à 38° chez les premiers, de 40 à 44° chez les seconds ; cela est possible parce qu’ils ont un revêtement mauvais conducteur de la chaleur, grâce à l’air enfermé dans le plumage et le pelage. Quand l’animal est à peu près nu, comme le Porc, la plupart des Mammifères aquatiques, l’Homme, des mécanismes internes de régulation ou une autre défense passive doivent suppléer celle qui fait défaut ; l’Homme non endurci se couvre de fourrure, de vêtements superposés entre lesquels s’intercalent des lames d’air ; avec une pelisse fourrée, un Homme perd cinq fois moins de chaleur que lorsqu’il est nu ; les animaux polaires ou de pays aux hivers rigoureux, comme l’Ours blanc, l’Oie, l’Eider ont un revêtement si efficace qu’ils peuvent supporter de très grands froids, tandis que la température d’un Mouton, après la tonte, s’abaisse sensiblement. Il est à peine utile de rappeler la régulation saisonnière des animaux à fourrure et leurs mues, ainsi que la présence d’une épaisse couche de graisse chez le Porc et les Cétacés.
Il y a donc partie liée, ou coaptation, entre le pelage- plumage et la fonction de régulation thermique ; dans nos climats, un Mammifère nu n’est pas viable ; c’est pour cela que l’Homme n’a pu apparaître que dans un climat tropical ; les races de Chiens nus ne se maintiennent que dans des pays très chauds (Mexique, Bolivie, etc.) ; le curieux Rongeur glabre d’Afrique, l’Heterocephalus glaber, est un fouisseur à mœurs de Taupe.
Puisque l’homéotherme ne peut se passer du revêtement protecteur, nous sommes forcés d’admettre que celui-ci est apparu avant le mécanisme de la régulation thermique (il y a un pelage dense chez l’Ornithorynque qui n’est encore qu’un médiocre homéotherme).
Mais alors il est bien difficile de ne pas attribuer à l’apparition du poil et de la plume la signification d’une préparation à l’acquisition de la température constante. La coaptation anatomo-physiologique exclut le hasard et suggère le plan.
On pourrait objecter qu’il y a des poils ailleurs que chez les Mammifères, par exemple chez les Insectes, les Araignées, les Crustacés, voire chez certains Annélides; mais il y a rarement un pelage; ce sont des poils sensitifs, ou des coussinets plantaires disposés pour la marche, ou des brosses revêtant des surfaces de roulement, ou des organes d’accrochage, ou encore des poils venimeux. II y a cependant des chenilles revêtues d’une véritable fourrure (chenilles de Macrothylacea rubi); elle est peut-être atélique3; elle est peut-être défensive. La nature a inventé plusieurs fois le poil pour des fins très variées.
Le pelage et le plumage, une fois créés, ont subi, bien entendu, tontes sortes d’avatars ; les hormones sexuelles ont exercé souvent leur action, modifiant les couleurs et poussant les formes jusqu’à l’hypertélie (Paon) ; en sens inverse, le poil a pu disparaître, remplacé par autre chose, en laissant des îlots spécialisés ou atéliques, comme les sourcils, les cheveux, la barbe, les poils de l’aisselle et de la région génitale.
Les callosités :
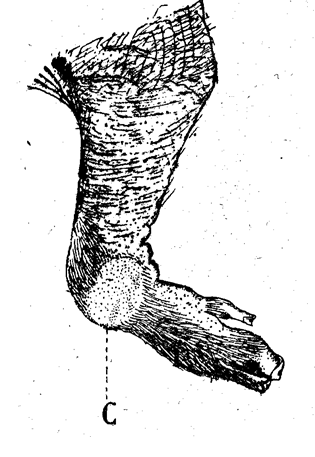
Fig.1. Patte antérieure gauche d’un fœtus un peu avant terme (Phacochœrus africanus)
C : callosité carpienne.
Une coaptation plus intéressante encore que la précédente est celle des callosités ; nous nous occuperons seulement de la callosité carpienne du Phacochère : ce Sanglier africain, aux formidables défenses, a coutume de fouiller constamment la terre avec l’extrémité charnue du museau et ses canines, pour déterrer des racines (c’est un grand ravageur des cultures de manioc) ; pour ce faire, il s’agenouille1 sur la région carpienne des membres antérieurs, se poussant en avant au moyen des pattes de derrière, appliquées sur le sol par toute la surface tarsopédieuse. Même en captivité et dans une cage étroite, il s’agenouille pour manger.
Quand on examine un Phacochère adulte, on voit de fortes callosités cornées sur les régions carpiennes et sous-calcanéennes, protégeant ces parties qui frottent rudement sur le sol. Or, les callosités carpiennes du Phacochère sont parfaitement visibles chez des embryons encore très jeunes (Leche, Cuénot, Anthony) (cf. fig. 1) ; ce sont des surfaces lisses, absolument glabres, dont l’épiderme épais et très plissé a une épaisseur triple de celle de l’épiderme banal avoisinant ; la couche cornée est bien formée et se desquame (fig. 2).
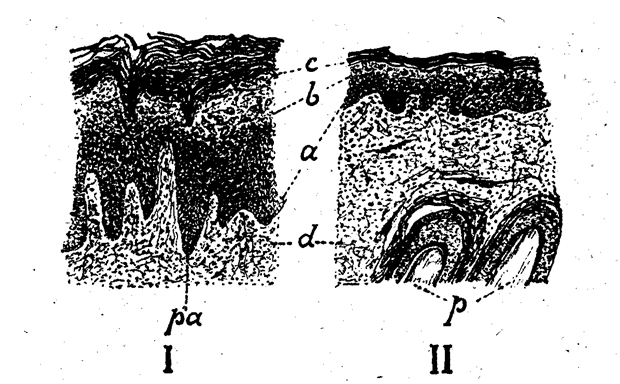
Fig. 2. — Coupe longitudinale de.la callosité carpienne d’un fœtus de Phacochère; les deux parties, représentées au même grossissement, montrent la grande différence d’épaisseur de l’épiderme ; la section I passe par la callosité ; la section II par la région normale qui l’entoure : a, couche germinative ; b, couche granuleuse ; c, couche cornée : d, derme ; p, poils coupés obliquement ; pa. papilles épidermiques très développées dans la région calleuse.
En somme, les callosités sont préparées chez l’embryon, alors que celui-ci, suspendu dans son amnios, n’a été en contact avec aucune surface dure. Cette même préparation est connue chez les embryons d’Autruche et de Nandou pour la callosité sternale, chez les jeunes Dromadaires et Chameaux pour leurs diverses surfaces d’appui (elles sont recouvertes de poils avant de devenir des callosités), chez les embryons de Singes de l’Ancien Monde pour les callosités fessières, chez le fœtus humain pour la plante du pied (épiderme déjà épaissi au 4ème mois de la vie intra-utérine), chez les fœtus de digitigrades pour les pelotes digitales des pattes, et d’une façon générale pour toutes les surfaces ou saillies qui seront plus tard en contact avec le sol. C’est maintenant un fait bien établi.
La callosité, ou plus exactement sa préparation fait partie de la totalité Phacochère, c’est-à-dire qu’elle est inscrite dans le patrimoine héréditaire ; elle existe en puissance dans l’œuf. C’est sans doute une acquisition récente, car on ne la trouve ni chez le Sanglier ni chez le Potamochère; elle est liée incontestablement à l’instinct qui porte le Phacochère à fouiller le sol en prenant une attitude particulière.
Deux hypothèses explicatives sont possibles : 1° la callosité organisée s’est formée, par une mutation de hasard, avant que l’animal ait pris l’habitude de s’agenouiller pour manger, et c’est parce qu’il avait déjà une callosité que l’instinct a pu aboutir à la manœuvre actuelle ; 2° l’instinct s’est développé d’abord. Par réaction au frottement, une callosité somatique s’est formée chez chaque individu, analogue aux cals professionnels déterminés chez l’Homme par le maniement des rames, de la bêche, etc. Après un nombre X de générations, la callosité acquise s’est inscrite, nous ne savons comment, dans le patrimoine héréditaire, si bien qu’elle est maintenant préparée chez le fœtus, avant toute excitation épidermique.
La première hypothèse, que l’on pourrait appeler préadaptationniste, n’est pas vraisemblable ; il faudrait admettre que le hasard aveugle ait produit l’organe très exactement à la place où le carpe sera (plus tard) en contact avec le sol; c’est un peu trop « cause-finalier ».
La seconde hypothèse, toute lamarckienne, repose sur la possibilité de la transmission héréditaire de caractères acquis par le soma; or, ce11e-ci est inconcevable et indémontrée dans l’état actuel de nos connaissances. Nous voici donc au pied du mur et toute issue paraît fermée ; voire ! Remarquons que la grande difficulté pour comprendre l’hérédité de la callosité, c’est la dissociation entre deux ordres de faits, l’un psychique, l’autre anatomique : l’instinct de s’agenouiller sur les carpes pour fouir, et la présence d’une callosité organisée. Mais pourquoi les dissocier ? On peut supposer qu’une variation unique du germen a eu un double effet :
1° en créant un instinct particulier, coordonné du reste au museau robuste, à la puissante armature dentaire du Phacochère et à la structure des pattes antérieures, 2° en inventant en même temps, et d’un coup, l’organe nécessaire à la satisfaction de l’instinct, c’est-à-dire la région préparée pour devenir une callosité quand il y aura frottement. On pourrait établir un parallèle entre le cas des Phacochères et celui des raboteurs de planchers qui ont inventé un coussin rembourré sur lequel ils s’agenouillent pour effectuer leur travail.
Je tiens l’exemple des callosités pour l’un des arguments les plus importants au point de vue d’une interprétation finaliste de la Vie ; après y avoir longtemps réfléchi et avoir retourné la question de toutes les façons, je ne vois aucune possibilité d’en donner une explication mécanistique : l’hypothèse lamarckiste, naturelle et séduisante, est au fond toute téléologique ; celle du mutationnisme l’est encore plus, puisqu’on voit poindre l’abhorrée cause finale ; quant à une thèse darwiniste, il est inutile d’en parler, car elle ne saurait être qu’une combinaison des deux précédentes.
1 Repris de Invention et Finalité en Biologie, Paris, Flammarion, 1941, pp. 160-166.
2 Membre de l’Académie de Sciences, naturaliste reconnu, Lucien Cuénot (1866-1951) était évolutionniste. C’est donc sous la contrainte irrémissible des faits qu’il admet ici la finalité manifestée chez les êtres vivants.
3 Ndlr. Du grec télos, fin, but, objectif. De là « a-télique ». Ici: qui n’a pas de finalité directe. Il est clair que c’est son évolutionnisme qui inspire à Cuénot l’idée que des organes résiduels peuvent avoir subsisté mais n’ont plus de finalité. En 1931, Alfred Wiedersheim avait établi une liste de 180 organes « vestigiaux » chez l’homme ! Le progrès des connaissances a permis de connaître la fonction ou les fonctions de tous ces organes.
1 Terme évidemment impropre, dont on se sert faute de mieux.