Partager la publication "La génétique de la domestication des céréales"
Par Pernès J
La génétique de la domestication des céréales1
Jean Pernès2
Résumé: Nous savons par la Genèse (1, 24 et 26) que Dieu a créé un petit nombre d’espèces animales destinées à être des compagnons et des serviteurs pour l’homme. Même si la Bible est muette sur les plantes domestiques, le fait est qu’un petit nombre d’espèces végétales sont indispensables à notre vie terrestre. Dans un article de La Recherche, l’auteur, généticien au CNRS, cherche à expliquer ce « syndrome de la domestication », le fait que des critères précis distinguent les variétés cultivées des variétés sauvages : maturité simultanée et attache solide des graines (pour la récolte sur pied), finesse de l’enveloppe (pour la digestibilité), germination rapide (pour les semis), etc. Si les gènes commandant ces qualités étaient répartis au hasard dans le génome, et si la domestication avait été le fruit d’une sélection dirigée (comme les agronomes la pratiquent aujourd’hui pour améliorer les variétés), il aurait fallu un très grand nombre de tentatives par essais et erreurs pour passer de la souche « sauvage » à une variété cultivable, ce dont l’histoire de l’agronomie n’a laissé aucune trace.
La génétique apporte aujourd’hui la clé de ce paradoxe : tous les gènes de la domestication sont portés par un seul et même chromosome.La première récolte a donc suffi pour pouvoir sélectionner et semer une variété domestique.
Enfermé dans sa vision scientiste du monde et de l’homme « préhistorique », le généticien Jean Pernès s’étonne donc devant l’inexplicable « révolution néolithique », qui aurait vu cette « invention » brusque de l’agriculture. Nous savons, quant à nous, que les êtres vivants, y compris les plantes cultivées, ont été créés par une Intelligence supérieure en vue d’une fin.
La solution est alors immédiate : il suffit de remplacer, dans cet article, « proto-agriculteurs du Néolithique » par : Adam et Caïn ! Une fois de plus, c’est la vision biblique du monde qui « colle » le mieux avec tous les faits connus.
Les plantes cultivées comme le blé, le maïs, le riz… sont issues de plantes sauvages. Bien souvent, l’aspect de la plante cultivée et celui de la plante sauvage dont elle dérive sont très différents. C’est que le processus de domestication a profondément altéré les caractères biologiques de la plante sauvage. Dans la plupart des cas, les céréales ont été domestiquées au sortir de la Préhistoire, il y a 10000 ans environ. Nos ancêtres du Néolithique avaient-ils donc un « savoir biologique » si étendu pour être capables de domestiquer des plantes en les transformant à ce point?
En fait, les travaux contemporains en science de l’hérédité montrent que la génétique de la domestication des céréales est, dans la plupart des cas, fort simple. C’est donc par des pratiques agronomiques familières que les premiers agriculteurs parvinrent à domestiquer les céréales.
Cela explique que la même plante a pu être domestiquée indépendamment en plusieurs endroits de la planète, comme cela s’est sans doute passé pour le riz.
À propos de cet épisode crucial de l’histoire de l’humanité, appelé « révolution néolithique », de nombreuses questions se posent : pourquoi l’homme, qui avait vécu durant les millénaires de la Préhistoire comme chasseur-cueilleur et nomade3, devint-il brusquement agriculteur et sédentaire ? Qu’est-ce qui l’a poussé à ce changement considérable de son mode de vie et comment cela s’est-il réalisé ? […]Mais se pose une tout autre question concernant les origines de l’agriculture est celle-ci : quel était le degré de savoir « biologique » ou de savoir-faire « agronomique » nécessaire pour domestiquer des plantes ? Pour dire les choses autrement, la domestication représente une « invention » ; il est donc fort probable que nos ancêtres du Néolithique aient pressenti qulques notions de génétique et maîtrisé certaines pratiques de « sélection artificielle » pour arriver à domestiquer les céréales.
Comment cela a-t-il été possible, alors que la connaissance scientifique de l’hérédité des caractères, la génétique précisément, ne date que de 1865, c’est-à-dire du moment où le moine autrichien Gregor Mendel découvrit les lois de la transmission héréditaire, en se livrant à des expériences d’hybridation sur les petits pois cultivés au monastère de Brno ?
La question du « savoir biologique » que possédaient nos ancêtres du Néolithique est encore plus étonnante lorsqu’on prend conscience du fait suivant : les plantes cultivées sont issues de plantes sauvages. Or, presque toujours, ces deux types de plantes diffèrent énormément par leur aspect. Le cas le plus spectaculaire est, sans doute, celui du maïs. Un pied de maïs n’a qu’une seule tige robuste qui ne porte qu’un ou deux grands épis (de 25 cm de longueur environ), sur lesquels se trouvent une vingtaine de rangées de graines.
Or le maïs dérive d’une espèce sauvage, appelée téosinte. Un plant de celle-ci est d’aspect bien différent de celui du maïs : il est composé d’une demi-douzaine ou plus de tiges frêles, portant de très nombreux petits épis (6 ou 7 cm de longueur environ) qui ne présentent qu’une seule rangée de graines ; celles-ci ont, de plus, un aspect triangulaire, fort différent de la graine quadrangulaire du maïs. Bref, les différences de morphologie entre maïs et téosinte sont telles que les botanistes ont, depuis le XVIIIe siècle, donné des noms scientifiques fort différents à ces deux plantes. Le maïs a été baptisé Zea mays et la téosinte Euchlena mexicana. En fait, maïs et téosinte paraissent tellement différents que beaucoup de botanistes admettaient encore, il y a une dizaine d’années, qu’il n’y avait aucun rapport direct entre ces deux plantes et que l’ancêtre sauvage du maïs restait inconnu.
[…] En définitive, tout cela donne l’impression que la domestication a changé considérablement l’aspect des plantes. Cela laisse penser qu’il a fallu un « savoir biologique » et un « savoir-faire agronomique » considérables pour obtenir les céréales cultivées à partir des formes sauvages. En réalité, des travaux de génétique menés ces quinze dernières années de par le monde sur les céréales, ont montré que la domestication de ces plantes avait, sans doute, suivi un processus assez simple.
En fait, la domestication n’a même été possible que parce que des particularités génétiques simples préexistaient déjà chez un certain nombre de plantes sauvages et qu’elles ont été mises à profit par des pratiques agronomiques également simples. Nous allons voir dans cet article de quelles particularités génétiques et de quelles pratiques agronomiques il s’agit. […]
Le syndrome de la domestication.
Une céréale cultivée présente un ensemble de caractères biologiques qui sont autant d’adaptations utiles à l’agriculteur : cet ensemble de caractères a reçu le nom de syndrome de la domestication. Par exemple, une céréale comme le blé, le maïs ou le mil, porte de gros épis, mais en petit nombre, arrivant à maturité à peu près simultanément. C’est là un caractère très utile à l’agriculteur : celui-ci désire en effet récolter le plus facilement possible la production de son champ en une ou deux fois, à la fin de la période de culture de la céréale.
Si l’on compare la plante cultivée à la plante sauvage dont elle dérive, on s’aperçoit que cette dernière ne se prête pas du tout à une telle facilité de récolte. Le mil sauvage, par exemple, présente en effet des plants ayant la forme de touffes buissonnantes, portant des centaines de petits épis arrivant à maturité de façon très échelonnée, alors que les plants du mil domestique comprennent au plus une dizaine d’épis portés par quelques grosses tiges, qui mûrissent quasi tous en même temps (Fig. 1).
En fait, le mil sauvage présente un certain nombre de caractères qui permettent une dispersion efficace des graines sur une grande période de temps et sur de grands espaces. Les graines sont, en effet, attachées sur l’épi par un pédicelle court qui se rompt facilement. Elles peuvent donc se détacher aisément au fur et à mesure de leur arrivée à maturité. Elles sont entourées de longues soies leur permettant d’être emportées au loin par le vent.
À l’inverse, les graines du mil domestique sont fermement attachées à l’épi par un pédicelle long et les soies sont pratiquement absentes. En définitive, on voit donc que la plante sauvage présente un ensemble de caractères assurant une dispersion maximale de ses graines, alors que la plante domestique présente un ensemble de caractères assurant précisément l’inverse (c’est-à-dire la non-dispersion) !


Fig. 1 : Épi de mil sauvage (à gauche) et mil domestique (à droite)
C’est que les « intérêts » de la reproduction de la plante sauvage et de l’agriculteur sont à l’exact opposé. Pour s’assurer les meilleures chances d’établir la génération suivante, la plante sauvage a « intérêt » à étaler la dispersion de ses graines mûres dans l’espace et dans le temps. Au contraire, s’il veut une récolte « rentable », l’agriculteur a intérêt à « rassembler » dans le minimum d’espace et de temps le maximum de graines matures.
Cette opposition des « intérêts » de la reproduction de la plante sauvage et de l’agriculteur se retrouve au niveau de nombreux autres caractères biologiques. Il en est ainsi, par exemple, pour la germination des graines. Chez les céréales sauvages, un bon étalement de la germination donne plus de chances à certaines plantules de rencontrer des conditions favorables.
Aussi, les graines des céréales sauvages présentent-elles de nombreux dispositifs qui tous retardent et rendent hétérogène le moment de la germination.
Les graines sont munies d’enveloppes épaisses qui devront être dégradées lentement avant de libérer la plantule ; il existe des mécanismes chimiques nombreux d’inhibition de la germination ; les besoins de croissance de la plantule, assurés par les substances stockées dans la graine, ne sont que progressivement satisfaits, car les substances de réserve sont des protéines, en proportion relativement élevée, c’est-à-dire des substances qui ne sont pas facilement mobilisables…
Au contraire, le cultivateur a besoin que les graines qu’il jette dans le champ labouré et préparé germent rapidement et toutes ensemble : de cette façon, elles pourront prendre de vitesse les mauvaises herbes et le champ sera donc uniformément couvert de céréales, ce qui facilitera naturellement la prochaine récolte. Donc, les graines des céréales cultivées doivent présenter peu de dispositifs retardant la germination ; les enveloppes des graines doivent être peu développées, et les réserves comprendre de fortes proportions de glucides.
Remarquons que les impératifs de la préparation culinaire vont aussi dans le sens de la diminution de la grosseur des enveloppes des graines et de l’élimination des soies.
En définitive, les plantes domestiques se distinguent donc des formes sauvages dont elles dérivent, par des caractères biologiques utiles à l’agriculteur pour la récolte, le semis ou la préparation culinaire. Remarquons enfin que plantes domestiques et plantes sauvages ne diffèrent pas du tout — contrairement à ce que l’on pourrait croire – quant au rendement productif de leur métabolisme.
La génétique des caractères utiles à l’agriculteur
La génétique des caractères utiles à l’agriculteur (c’est-à-dire du syndrome de domestication) a été particulièrement étudiée dans les années 1970, en ce qui concerne le maïs, par J. R. Harlan et J. M. J.de Wet, de l’université d’Urbana, Illinois (États-Unis) et par W. Galinat, également aux États-Unis. Nous avons nous-mêmes, avec notre équipe, éclairci, durant cette même période, la question du contrôle génétique du « syndrome de domestication » chez le mil.
Tout d’abord, il faut savoir que, malgré leur morphologie fort différente et leur désignation latine distincte, le mil sauvage et le mil domestique appartiennent à la même espèce biologique : leurs hybrides sont entièrement fertiles. D’ailleurs, dans les localités africaines où mil sauvage et mil domestique coexistent, des croisements spontanés ont lieu régulièrement. Les cultivateurs africains ont toujours reconnu les hybrides dans leurs champs, notamment au fait qu’ils possèdent des caractères biologiques impropres à la culture : les graines se détachent avant que l’ensemble de l’épi soit mûr ; elles sont munies de grandes soies et de grosses enveloppes ; les différentes tiges ne fleurissent pas en même temps… Chaque ethnie a même donné un nom particulier à ces hybrides : ndoul en wolof, chibra en haoussa.
Nous avons pu faire une analyse génétique de la descendance de ces hybrides. En premier lieu on constate que, parmi les plantes issues de ces hybrides, certaines sont absolument indistinguables d’une plante sauvage. Autrement dit, les caractères des parents réapparaissent intégralement dans la descendance des hybrides.
Cela signifie que les lois de Mendel sont applicables à l’ensemble du syndrome de domestication. Si l’on prend un à un chacun des caractères du syndrome de domestication du mil, on s’aperçoit qu’il se présente, sur le plan génétique, comme une alternative à un caractère de la plante sauvage. Par exemple, le long pédicelle (attachant fermement la graine sur l’épi), qui s’observe chez le mil domestique, est remplacé par un pédicelle ultra-court (rendant la graine facilement détachable de l’épi), chez le mil sauvage. L’analyse génétique montre que le caractère long ou court du pédicelle sont les deux versions possibles d’un même gène. Il en va de même pour les autres caractères du syndrome de domestication : un seul gène à deux versions différentes détermine si les graines sont enveloppées ou non ; si les soies sont nombreuses ou non ; si la graine tombe ou non lorsqu’elle est mûre. Pour la longueur des soies, les choses sont un peu plus complexes : deux gènes interviennent en même temps.
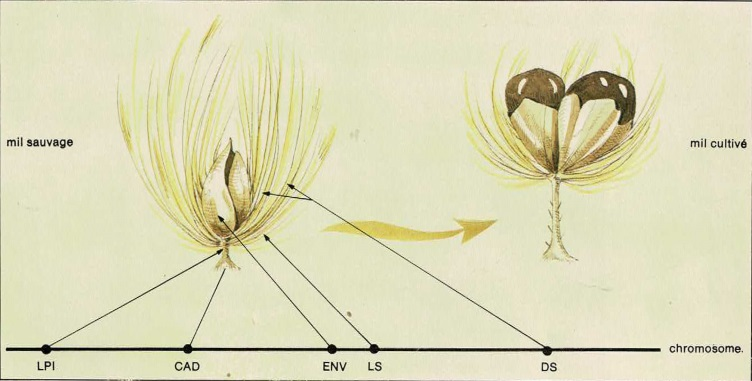
Fig. 2 : Gènes liés à la domestication chez le mil. LPI (longueur du pédicelle), CAD (brisure du pédicelle), ENV (forme des enveloppes de la graine), LS et DS (longueur et densité des soies). Ces 5 gènes de la domestication, proches les uns des autres sur le même chromosome (l’axe horizontal mesure la distance en centimorgans), seront transmis « en bloc » à chaque génération.
En définitive, on voit donc que le syndrome de domestication consiste, pour un plant de mil, dans le choix d’une des versions du gène contrôlant chacun des caractères. Finalement, l’ensemble du syndrome de domestication chez le mil paraît se comporter comme une version d’une seule grande unité génétique globale, constituée par l’ensemble des caractères considérés.
C’est ce que notre équipe a montré par des analyses génétiques plus précises (C. Rey-Herme, 19814; O. Niangado, 19815). Les différents gènes, déterminant les caractères (longueur du pédicelle, caducité de l’épillet à maturité, enveloppement de la graine, densité des soies) sont portés par le même chromosome (Fig. 3).
La version, utile à l’agriculteur, de chacun de ces gènes peut donc être transmise à chaque génération, solidairement avec les versions utiles de chacun des autres gènes participant au syndrome de domestication. C’est ce qui explique que, malgré la multiplicité des gènes mis en jeu dans le syndrome de domestication, le mil cultivé conserve, d’une génération à l’autre, avec l’assistance d’une épuration vigilante faite par le paysan, l’ensemble des caractères utiles à l’agriculteur. Autrement dit, la forme cultivée du mil peut jouir d’une stabilité génétique. C’est aussi ce qui explique que, dans la descendance des hybrides du mil sauvage et du mil domestique, certaines plantes retournent « en bloc »… au type cultivé… Ce qui est vrai pour le mil, l’est également pour le maïs. Chez cette céréale aussi, les caractères utiles à l’agriculteur résultent de versions particulières de gènes localisés sur le même chromosome, et se transmettant solidairement «en bloc ». Dans le cas du maïs ce bloc est même bien plus étroitement soudé que chez le mil.
Finalement, l’analyse génétique montre encore que les plantes cultivées et leur forme sauvage ne diffèrent pratiquement que par leurs gènes relatifs au syndrome de domestication. Leurs autres gènes sont, dans leur immense majorité, identiques. Cela veut dire qu’au bout du compte, un assez petit nombre de gènes — une dizaine environ — ont été soumis à un choix déterminé durant le processus de la domestication.
Les actions des premiers agriculteurs.
Peut-on maintenant, sur la base de ces données génétiques, comprendre comment a pu se faire, techniquement, le processus de domestication du mil, par exemple ?
Il est généralement admis aujourd’hui que l’agriculture a commencé par la récolte périodique d’épis de certaines plantes sauvages. De nos jours encore, on peut voir se dérouler certaines phases de proto-agriculture concernant certaines céréales. Ainsi, aux abords du lac Tchad, les habitants de la région récoltent quotidiennement les épis de populations sauvages de riz (Fig. 3) : on peut facilement vérifier que seuls tombent dans les paniers les graines arrivées à maturité et donc capables de se détacher.
Or on peut constater aussi que les récolteurs ont mis au point certaines méthodes afin de limiter la fréquence de leur passage, tout en augmentant le nombre de graines recueillies à chaque fois. Ces méthodes consistent, par exemple, à ligaturer les épis, de telle façon que les graines restent emmêlées par leurs barbes et empêchées ainsi de tomber au sol…

Fig. 3. Récolte du riz sauvage au Tchad, faite par les femmes et les enfants armés de paniers6.
Il est donc évident que les récolteurs, du fait même de leur activité de cueillette, sont prêts à saisir toute occasion qui leur est offerte pour atteindre l’objectif du maximum de graines avec le minimum de travail. Or, comme nous l’avons vu ci-dessus, les caractères tels que la longueur du pédicelle, etc., qui retardent la chute des graines, préexistent naturellement dans les populations sauvages. Il est évident, d’après l’exemple ci-dessus du riz sauvage récolté au Tchad, que les récolteurs du Néolithique devaient spécialement cueillir les épis portant les graines qui arrivaient en plus grand nombre à maturité et restaient le plus longtemps regroupées ensemble sur l’épi… Il leur restait alors à faire la découverte simple, mais capitale, que les graines semées redonnent des plantes.
À partir du moment où les proto-agriculteurs se mirent à semer les graines qu’ils avaient récoltées et cessèrent de se borner à récolter simplement les épis des plants sauvages, la sélection artificielle des caractères désirés s’ensuivit presque automatiquement.
En effet, en cueillant préférentiellement les graines arrivant à maturité toutes ensemble sur l’épi et restant plus sur l’épi, ils réalisaient une sélection de graines porteuses d’une version des gènes appartenant au syndrome de la domestication. Et en semant les graines qu’ils avaient ainsi récoltées, ils favorisaient forcément la multiplication des plantes présentant les versions génétiques particulières du syndrome de domestication.
Autrement dit, les caractéristiques héréditaires du syndrome de domestication sont suffisamment peu nombreuses et simples pour que des personnes en permanence préoccupées par leurs activités de récolte aient pu réaliser, même sans un « savoir biologique » étendu, la sélection des plantes cultivées. Cela est d’autant plus probable que la majorité des caractéristiques du syndrome de domestication ont dû être sélectionnées progressivement « autour » de la propriété initiale la plus importante, la perte de la caducité des graines.
Éliminer les hybrides…
Un problème se pose cependant : dans les zones géographiques où ont été domestiquées les céréales, les plantes cultivées coexistaient avec les plantes sauvages. Des champs de plantes cultivées pouvaient fort bien être environnés, même de façon dispersée, par les formes sauvages de la céréale. Dès lors, des plantes hybrides avaient sans doute tendance à se former à chaque génération, menaçant ainsi de faire disparaître le type génétique de la plante cultivée, et rendant ainsi malaisé le travail de l’agriculteur. Comment les premiers paysans arrivèrent-ils à maîtriser ce problème ?
Là encore, ils mirent à profit certaines particularités génétiques simples des céréales. En effet, pour plusieurs céréales tels le sorgho, le blé, le millet, l’orge, le riz, les formes sauvages se reproduisent préférentiellement par autofécondation, c’est-à-dire la fécondation de chaque plante par son propre pollen. Les formes cultivées du sorgho, du blé, du millet, de l’orge et du riz se reproduisent elles aussi préférentiellement par autofécondation. Dans ces conditions, on comprend bien qu’il y a peu de chances que formes sauvages et formes cultivées s’hybrident, là où elles coexistent !
Pour d’autres céréales cependant, tels le maïs et le mil, le mode de reproduction est préférentiellement croisé : le pollen d’une plante va féconder les ovules d’une autre plante. Dans ce cas, c’est le regroupement des gènes du syndrome de domestication sur un même chromosome qui va éviter au type cultivé de disparaître rapidement. En effet, supposons que le syndrome de domestication soit constitué de dix gènes indépendants, c’est-à-dire situés sur dix chromosomes différents. Supposons encore que les plantes cultivées aient été fécondées par le pollen des plantes sauvages. Des hybrides en résultent. Ces hybrides auront des descendants : mais seulement chez un sur un million de ces descendants, les dix gènes du syndrome de domestication se retrouveront par hasard réunis. Donc, la restitution du type cultivé à partir des hybrides n’a qu’une très faible probabilité de se produire. Dans ce cas, même si les populations sauvages n’assurent pas un nombre très élevé de croisements avec les populations cultivées, le type cultivé risque néanmoins de disparaitre assez rapidement.
Au contraire, si les dix gènes du syndrome de domestication sont portés en bloc sur le même chromosome, ils se transmettent comme s’ils étaient presque un seul gène. La probabilité de voir resurgir le type cultivé dans la descendance des hybrides entre plant sauvage et cultivé est élevée : 25 % des descendants de croisements entre de tels hybrides sont de type cultivé, s’il n’y a réellement que l’équivalent d’un seul gène en jeu ; mais 50 % des descendants de croisements entre hybrides et plants cultivés sont de type cultivé. Dans les conditions où le système génétique tend à préserver le type cultivé, l’homme n’a que peu de mal à compléter l’action de la nature. Dans les régions où les champs de mil cultivé peuvent être « contaminés » par le pollen du mil sauvage, le paysan peut, sans être débordé, faire la chasse aux hybrides dans son champ, pour les détruire au moment de la floraison, avant donc qu’ils ne donnent des graines à leur tour. L’homme, par sa vigilance, préserve la stabilité génétique de ses plantes cultivées telles que le mil ou le maïs, stabilité déjà promue par le système génétique particulier de ces espèces : la cohésion en bloc des gènes du syndrome de domestication.
Cette vigilance, qui fut nécessaire aux premiers agriculteurs qui tentèrent la domestication du mil, l’est encore aujourd’hui7. En effet, en Afrique, encore actuellement, le mil cultivé par exemple est en permanence en situation de contact avec les populations de mil sauvage. Les cultivateurs traditionnels africains savent sauvegarder leur mil par la destruction vigilante des hybrides si possible dès la germination des plantules. À l’inverse, des ingénieurs agronomes d’un centre de recherche en Inde, qui se proposaient en 1980, de « conserver les ressources génétiques » du mil, eurent la mésaventure de voir se dégrader leurs variétés de mil : ils avaient « oublié » de détruire les hybrides qui s’étaient glissés dans leurs collections !
En définitive, il semble bien que la domestication des céréales n’ait été possible que pour des plantes qui présentaient des particularités génétiques telles que le regroupement des gènes favorables sur le même chromosome ou des particularités reproductives telles que l’autofécondation. Il n’est pas impossible que des tentatives de domestication de céréales, autres que celles actuellement connues, aient eu lieu dans le passé. Simplement, de telles tentatives ont échoué parce que de telles plantes n’avaient pas de dispositions génétiques ou reproductives qui favorisaient le maintien d’un type cultivé. Cela a peut-être été le cas pour la graminée appelée Cenchrus, qui pousse actuellement au Sahel. Des découvertes archéologiques ont montré que cette céréale fut l’objet d’une récolte à des fins alimentaires, il y a plusieurs milliers d’années. Mais cette plante n’a jamais été cultivée. Sans doute, sa génétique ne s’y prêtait pas.
… mais ne pas les éliminer tous !
Pour finir, en ce qui concerne la destruction des hybrides, il faut ajouter que l’action des paysans ne permet jamais leur élimination totale. Dans les aires d’origine des céréales, il reste toujours, dans les champs cultivés, une toute petite proportion d’hybrides. Or cette petite proportion n’est pas nuisible, au contraire. Ces hybrides représentent une sorte de « cheval de Troie » par lequel des qualités génétiques nouvelles, provenant des populations sauvages, peuvent se répandre dans les populations cultivées. Ceci peut être extrêmement utile à l’agriculteur : si une maladie se répand dans ses cultures, il aura besoin de nouvelles variétés dotées de gènes de résistance à cette maladie. Or ces gènes ont de fortes chances d’exister dans les populations sauvages.
Du Néolithique à aujourd’hui, les paysans ont su tirer parti de ces hybrides spontanés entre populations sauvages et cultivées, pour mettre au point de nouvelles variétés de leurs plantes cultivées, résistantes à de nouvelles maladies ou dotées d’autres caractères nouveaux, jugés désirables.
Encore aujourd’hui, les paysans des diverses régions d’origine des céréales savent qu’il ne faut pas détruire la totalité des hybrides entre populations sauvages et cultivées. C’est le cas pour les cultivateurs du mil dans la région du Sahel (du Tchad au Sénégal), du riz dans le delta du Niger, du millet dans le Chang-Si en Chine, du blé en Iran, Afghanistan ou Palestine…
L’amélioration moderne des plantes, au moyen des techniques sophistiquées de la génétique, est d’ailleurs, elle aussi, tributaire de la diversité génétique entretenue sans cesse dans les populations naturelles (La Recherche, n° 38, p. 867, octobre 1973). Cela montre en retour combien il est nécessaire de maintenir les ressources génétiques que constituent ces populations et que l’agriculteur traditionnel gère intuitivement quand il surveille son champ et choisit ses semences. Si l’on veut éviter des catastrophes agronomiques dans le futur, il faut soigneusement préserver tant les populations des formes sauvages que les variétés traditionnelles (La Recherche, n° 84, p. 1 067, décembre 1977).
Des domestications à plusieurs reprises.
Ainsi, malgré les grandes différences de morphologie entre plantes cultivées et plantes sauvages, la domestication a été obtenue au moyen de pratiques agronomiques simples. Biologiquement, rien ne s’oppose donc à ce qu’à partir de populations sauvages d’une espèce donnée, des hommes aient réalisé indépendamment, en des lieux et des époques différents, la domestication d’une même céréale.
Aussi, si le biologiste russe Vavilov avait, entre les deux guerres mondiales, reconnu des centres géographiques d’origine pour un certain nombre de céréales, le biologiste américain J. R. Harlan introduisit, dans les années 1960, le concept de « non-centre » pour l’origine géographique de certaines d’entre elles (La Recherche, n° 29, décembre 1972, p. 1 035) : il s’agit d’aires très étendues où la domestication de certaines plantes a pu se faire sans que cela résulte d’une diffusion à partir d’un lieu d’origine géographiquement restreint. Autrement dit, au sein de ces vastes « non-centres » géographiques, la domestication d’une céréale donnée a pu se faire indépendamment en plusieurs endroits, alors que Vavilov imaginait une origine unique et forcément restreinte à partir de laquelle la diffusion se faisait par migration des plantes et des sociétés. Par exemple, toute la zone du Sahel, au sud du Sahara, serait un vaste « non-centre » pour la domestication des mils et des sorghos.
[…] En conclusion, les analyses génétiques permettent de ramener à de justes proportions l’importance de la différenciation génétique due à la domestication : pour spectaculaire qu’elle soit, elle ne concerne cependant qu’un nombre limité de structures génétiques. En conséquence, ce processus ne doit être ni très lent ni exceptionnel, à partir du moment où des situations sociales existent qui tendent à attirer l’attention vers les caractères dont la conservation et l’accumulation conduisent à l’aspect ou aux propriétés dits « du syndrome de domestication ». Il est alors vraisemblable que plusieurs domestications aient eu lieu indépendamment pour une même céréale et avant même que des sociétés aient acquis l’agriculture comme système général d’approvisionnement. À l’époque de la genèse de l’agriculture, les céréales ainsi prédomestiquées auraient été distribuées et cultivées plus largement. Les variétés ont pu être modelées pour s’adapter à des milieux différents grâce, tout particulièrement, aux flux de gènes issus des formes sauvages, sans toutefois perdre leurs structures de plantes cultivées. Cette dernière situation d’équilibre entre échanges génétiques spontanés et sélection par le paysan reste encore remarquablement vivante et organisée dans de nombreuses zones d’agriculture traditionnelle.
Les conséquences pratiques des analyses génétiques sont importantes ; elles permettent d’envisager des stratégies d’exploitation des formes sauvages pour créer des variétés mettant plus largement à contribution les potentiels adaptatifs des populations sauvages. L’organisation des ressources génétiques peut être rendue dynamique en simulant les pratiques des paysans des centres géographiques d’origine pour construire des populations variétales intégrant régulièrement les polymorphismes adaptatifs des populations sauvages spontanées. Celles-ci ont été confrontées depuis les temps géologiques à la diversité des adversités biologiques et physiques et ont accumulé une « mémoire génétique adaptative » considérable. La sélection faite par les paysans des centres d’origine s’est réalisée, fait remarquable, sans appauvrissement inutile de la variabilité génétique grâce à ces flux de gènes entretenus à partir de ces immenses réservoirs naturels spontanés. Ceux-ci sont hélas en cours de disparition, du fait des remaniements importants imposés par l’agronomie moderne.
Cette protection des formes sauvages et de leur connexion avec les formes cultivées est à décider de façon urgente ; elle suppose une organisation politique complexe car pour des plantes telles que le blé, l’orge, le maïs, les grands pays producteurs (États-Unis, Canada, France) ne sont pas les pays d’origine et il ne s’y trouve pas de populations sauvages. Pour le riz, les grands producteurs (Chine, Japon) ont fait disparaître chez eux la quasi-totalité des populations de riz sauvage ! La survie des céréales dans les grands pays producteurs dépend maintenant de la sagacité de la conservation des ressources génétiques des formes traditionnelles et sauvages dans d’autres pays dont l’agronomie n’est pas encore trop modernisée, dans le sens des abus technologiques destructeurs de diversité.
1 Reprise partielle de La Recherche n°146, juillet-août 1983, p. 910-919. Le lecteur pourra s’y reporter pour les références aux revues savantes. Souligné par nous.
2 Professeur de génétique à l’université de Paris XI et directeur du laboratoire de génétique et de physiologie du développement des plantes au CNRS, J. Pernès fut également consultant auprès de diverses institutions internationales pour l’organisation de banques de gènes en milieux tropicaux.
3 Ndlr. L’Auteur est ici victime de la confusion entre « primitif » et « sauvage ». En réalité les peuples dits primitifs ne sont pas comme les vestiges des premières sociétés humaines, mais des rameaux maintenus par les circonstances à l’écart des civilisations dont ils provenaient. Lire à ce sujet les divers articles publiés dans Le Cep n°70, février 2015 : « Les sauvages sont-ils des primitifs ? »
4 Thèse de 3e cycle, Orsay, 1981.
5 Id.
6 Ndlr. Les grains récoltés par cette technique peu sophistiquée – ceux qui, à maturité, se détachent par secouage – donneront en majorité, s’ils sont semés, une variété d’emblée domestique.
7 Il en va toujours de même lorsqu’un paysan cultive un « contrat de semence » pour un sélectionneur : certaines variétés ou espèces autres sont éliminées du champ au cours de la culture, par exemple l’orge ou le seigle dans le blé, car les grains ne pourraient plus être bien séparés par un triage mécanique après récolte.